Wheat individual grain-size variance originates from crop development and from specific genetic determinism
Texte intégral
Figure


Documents relatifs
No significant difference in the behaviour of coarse and fine-grained calcium carbonate was observed (Figure 8), although we note that the range of grain-size studied here is
Table 6.3 shows recognition performance for various synthesis configurations: natural speech spoken by human, manually determined costs (baseline), automatically deter- mined
The cells were two liter beakers having a lead sheet cylinder as cathode, duriron anodes, and a porous pot containing the anode and anode solution.. The porous pot is
Marti, “ Empirical rheology and pasting properties of soft-textured durum wheat (Triticum turgidum ssp. durum) and hard-textured common wheat (T. aestivum) ,” Journal of Cereal
So the highly densified ferrites with large grain can be obtained by the coprecipitated powder, addition of BaO.B,O, and sintering by the vacuum sintering
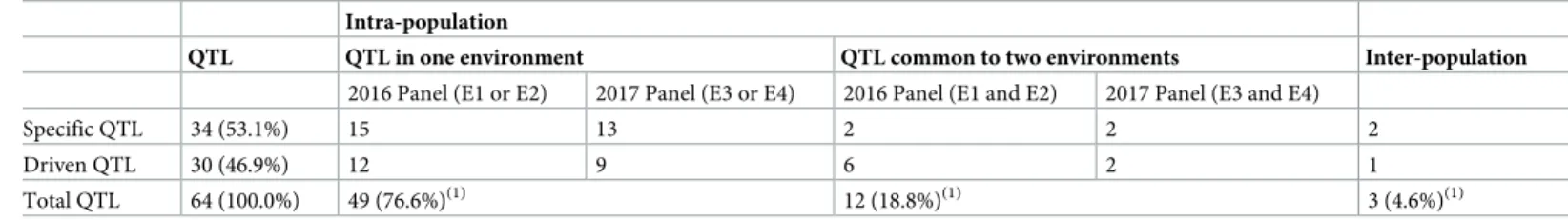
The predictions provided by the three models for the joint effects of population size and selection intensity on the evolution of genetic variance were compared. Figure
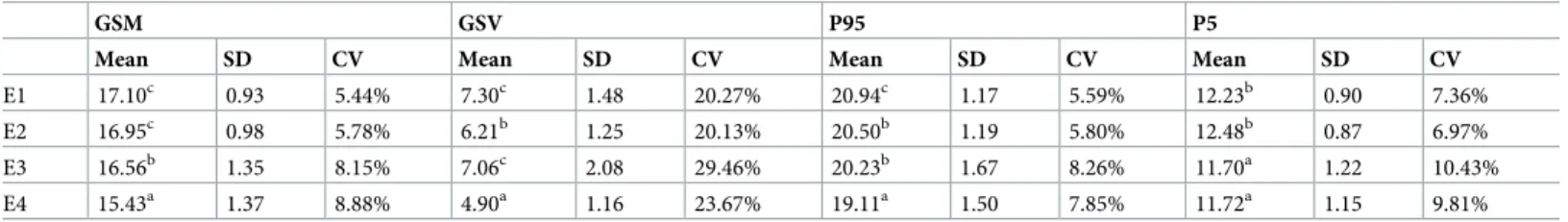
Statistics are calculated using arithmetic and geometric Method of Moments (micrometer) and us- ing logarithmic Folk and Ward (1957) Method (phi scale): mean,
Thus large grain sizes, which induce microcracks due to anisotropic volume changes on cooling from the sintering temperature, result in a decrease in jc with a
