HAL Id: hal-02262584
https://hal-amu.archives-ouvertes.fr/hal-02262584
Submitted on 11 Sep 2019
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
periodontitis
Sarah Benabdelkader, Julien Andreani, Alexis Gillet, Elodie Terrer, Marion
Pignoly, Herve Chaudet, Gérard Aboudharam, Bernard La Scola
To cite this version:
Sarah Benabdelkader, Julien Andreani, Alexis Gillet, Elodie Terrer, Marion Pignoly, et al.. Specific clones of Trichomonas tenax are associated with periodontitis. PLoS ONE, Public Library of Science, 2019, 14 (3), pp.e0213338. �10.1371/journal.pone.0213338�. �hal-02262584�
Specific clones of Trichomonas tenax are
associated with periodontitis
Sarah Benabdelkader1☯, Julien Andreani1☯, Alexis Gillet2, Elodie Terrer1,2,
Marion Pignoly2, Herve Chaudet1, Gerard Aboudharam1,2, Bernard La ScolaID1*
1 Aix-Marseille Universite´ UM63, Institut de Recherche pour le De´veloppement IRD, Assistance Publique– Hoˆpitaux de Marseille (AP-HM), Microbes, Evolution, Phylogeny and Infection (MEΦI), Institut Hospitalo-Universitaire (IHU) - Me´diterrane´e Infection, Marseille, France, 2 UFR Odontologie, Aix-Marseille Universite´ , Marseille, France
☯These authors contributed equally to this work.
Abstract
Trichomonas tenax, an anaerobic protist difficult to cultivate with an unreliable molecular
identification, has been suspected of involvement in periodontitis, a multifactorial inflamma-tory dental disease affecting the soft tissue and bone of periodontium. A cohort of 106 peri-odontitis patients classified by stages of severity and 85 healthy adult control patients was constituted. An efficient culture protocol, a new identification tool by real-time qPCR of T.
tenax and a Multi-Locus Sequence Typing system (MLST) based on T. tenax NIH4
refer-ence strain were created. Fifty-three strains of Trichomonas sp. were obtained from peri-odontal samples. 37/106 (34.90%) T. tenax from patients with periodontitis and 16/85 (18.80%˚) T. tenax from control patients were detected by culture (p = 0.018). Sixty of the 191 samples were tested positive for T. tenax by qPCR, 24/85 (28%) controls and 36/106 (34%) periodontitis patients (p = 0.089). By combining both results, 45/106 (42.5%) patients were positive by culture and/or PCR, as compared to 24/85 (28.2%) controls (p = 0.042). A link was established between the carriage in patients of Trichomonas tenax and the severity of the disease. Genotyping demonstrates the presence of strain diversity with three major different clusters and a relation between disease strains and the periodontitis severity (p<0.05). More frequently detected in periodontal cases, T. tenax is likely to be related to the onset or/and evolution of periodontal diseases.
Introduction
Periodontal disease is a widespread oral disease affecting adults and younger people, character-ized by an inflammatory reaction that affects periodontium tissue [1]. A new classification published in 2018 based on description (localized or generalized), severity and complexity of management divides periodontitis into 4 stages, including initial periodontitis (I), moderate periodontitis (II), severe periodontitis with potential for additional tooth loss (III) and advanced periodontitis with extensive tooth loss and potential for loss of dentition (IV) [2]. According to this classification, periodontitis is also graded in 3 levels estimated with direct or
a1111111111 a1111111111 a1111111111 a1111111111 a1111111111 OPEN ACCESS
Citation: Benabdelkader S, Andreani J, Gillet A,
Terrer E, Pignoly M, Chaudet H, et al. (2019) Specific clones of Trichomonas tenax are associated with periodontitis. PLoS ONE 14(3): e0213338.https://doi.org/10.1371/journal. pone.0213338
Editor: Peter Eickholz, Klinikum der Johann
Wolfgang Goethe-Universitat Frankfurt Klinik fur Nuklearmedizin, GERMANY
Received: October 3, 2018 Accepted: February 20, 2019 Published: March 11, 2019
Copyright:© 2019 Benabdelkader et al. This is an open access article distributed under the terms of theCreative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability Statement: The T. tenax NIH4
genome sequence was deposited on the EMBL-EBI website (Bioproject: PRJEB22701 and whole contigs under accession numbers OCTD01000001-OCTD01004161).
Funding: This work was supported by a grant from
the French State managed by the National Research Agency under “Investissements d’avenir (Investments for the Future)” program with the reference ANR-10-IAHU-03
(Me´diterrane´e-indirect evidence of progression rate: slow (A), moderate (B) and rapid (C). Periodontal dis-ease is characterized by receding gums, alveolar bone destruction, loss of dental junctions asso-ciated with the apparition of periodontal pockets, and in some forms, dental calculus deposits. This promotes the establishment of an anaerobic microenvironment that allows the growth of anaerobic microorganisms [3]. The immunological process initiates the migration of microor-ganisms into tissues and disrupts the immune response, causing the periodontium to resorb [4,5]. Some host risk factors have now been clearly identified, including smoking [6] and dia-betes mellitus [7], but other genetic factors require further study [8].
The human oral cavity contains an abundant and polymorphic microbiota. A set of bacte-rial complexes living in subgingival plaque has been identified by Socransky et al; with the ini-tial establishment of Streptococcus spp. and their disappearance, multiple complexes defined by green purple or yellow colors have been described but only the orange and red complexes have been suspected of being associated with the development of periodontitis [3]. Metage-nomic studies confirm the association of certain bacterial species found in the orange complex, such asPrevotella intermedia, Prevotella nigrescens and Fusobacterium nucleatum [9,10]. But also the strong association of the disease with the three bacteria described in the red complex: Pophyromonas gingivalis, Tannerella forsythia and Treponema denticola are also found in the periodontal pockets by metagenomic recent studies[11,12] and new species involved have recently been identified [13]. Nevertheless, the “red complex” theory remains debatable due to its isolation in healthy controls (30%, 3/10) [14]. An increase in lytic phages in pathological sit-uations also disturbs the periodontal-associated bacteria present [15]. Similarly, meta-tran-scriptomics analysis showed that the transcription of bacterial virulence factors increased in patients with periodontitis compared to healthy individuals [16]. Viral etiology has also been suggested as being involved in the development of periodontitis, and different viruses have been involved including Herpesviruses (HSV-1), Cytomegalovirus (CMV) and Epstein-Barr virus (EBV) [17].
Since the 1980s, the implication of eukaryotes in periodontal disease has also been proposed [18], such as the protistsTrichomonas tenax and Entamoeba gingivalis [19–21] and yeasts, such asCandida sp. [22]. However, until now, studies focusing on the association between pro-tists and periodontal disease did not use groups of healthy controls to differentiate between abnormal proliferation and natural colonization [20]. Like its neighboring species, Trichomo-nas vaginalis, the vaginitis-inducing pathogen, T. tenax belongs to the Parabasalia phylum and to theTrichomonadidae family [23].Trichomonas tenax can ingest bacteria and various parti-cles by phagocytosis necessary for their development[24]. Initially identified as an harmless commensal [18], then known as a zoonotic parasite [25], this microorganism was detected in the periodontal pockets using mostly optical microscopy [26,27], with an occurrence in patients ranging from 0 to 94.1% depending on the country and the detection procedure [21]. T. tenax has occasionally been isolated in cases of the salivary glands, lymph nodes or respira-tory tract infections [28][29][30]. Recently,T. tenax was found to be significantly more preva-lent in patients with Down syndrome combined with periodontal lesions (14/52), using 18S rRNA gene PCR in comparison to control patients (5/52), with a non-significant difference in plaque indexes between the two groups [31].
In this cohort study, we sought to estimate the prevalence ofT. tenax and establish a poten-tial link with the periodontitis severity. We investigated the presence ofT. tenax in periodonti-tis as compared to the healthy controls using culture and quantitative molecular detection systems. We also used a genome-based system of strain typing to investigate the possibility that a clone or a group of clones ofT. tenax with particular pathogenicity are involved in periodontitis.
Infection) and by Re´gion Provence-Alpes-Coˆte d’Azur and European funding FEDER PRIMI. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Competing interests: The authors have declared
Materials and methods
Clinical sample collection and treatment
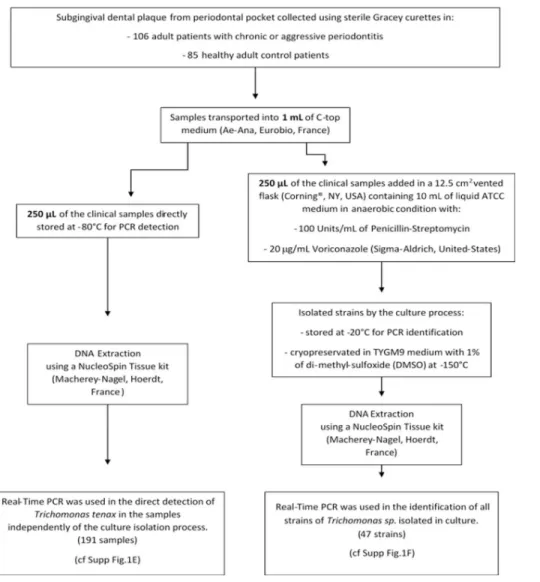
This study was carried out in accordance with the recommendations and approved by the clin-ical research ethics committee, IFR 48, Aix-Marseille University (protocol N˚ 2016–011). All subjects gave a written informed consent in accordance with the Declaration of Helsinki. One hundred and ninety-one adults were prospectively enrolled in this cohort, 106 adult patients with periodontitis and 85 healthy adult control patients, (S1 Table). The study took place in the Odontology Department of the Hospital La Timone, Marseille, France, between January 2015 and June 2016. The 106 patients with periodontitis could be separated in three classes of severity: 19 patients with mild periodontitis (M), 27 with moderate periodontitis (Mo) and 60 with severe periodontitis (S) according to a previously reported scale using various criteria evaluation: size of the probing depth and the attachment loss for each patient [32]. Smoking status was also collected (S1 Table). Subgingival dental plaque samples were collected from multiple periodontal pocket and combined into a single tube per patient for the group test and the same method was used for healthy gingival sites in the control group. The sample was col-lected using a sterile Gracey curette (HuFriedy, Rotterdam, Netherlands) and transported into 1 mL of transport medium (C-top Ae-Ana, Eurobio, France). The samples were analyzed according to the detailed protocol inFig 1.
Isolation of
Trichomonas tenax
Twelve-and-a-half cm2vented flasks (Corning, NY, USA) containing 10 mL of liquid ATCC: 1171 TYGM-9 medium without rice starch were used to inoculate 250μL of the clinical sam-ples. The medium was supplemented with 100 Units/mL of Penicillin-Streptomycin Gibco (ThermoFisher, MA, USA) and 20μg/mL Voriconazole (Sigma-Aldrich, United-States) to retard the bacterial and fungi growth that may interfere with theT. tenax development. Flasks were then incubated under anaerobic conditions using Anaerogen generators (ThermoFisher, MA, USA) at 35˚C. Growth was observed by optical microscopy examination. For the strains cryopreservation, cultures were centrifuged at 720 x g for 10 minutes. The final pellet was sus-pended in 1 mL of TYGM9 medium containing 1% of di-methyl-sulfoxide (DMSO) and placed in a Nunc cryotube, maintained at -80˚C for between 15 and 20 hours before being placed inside a -150˚C freezer.
Molecular identification and typing of isolates
Primer design. Based on the analysis of Maliket al.[33], we designed specific primers in the
3,048 bp sequence of the RNA polymerase IIrpb1 gene available on the NCBI website (accession number: HM016234.1 forT. tenax strain NIH4). Primers were designed using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) with standard parameters (Fig 1F).
Genome sequencing ofTrichomonas tenax. Briefly, 15 vented Corning 75cm2(NY,
USA) flasks containing LYI medium ofT. tenax NIH4 reference strain (ATCC number 30207) were pelleted at 2000 g for 15 minutes, rinsed twice with the same centrifugation parameters in Page’s amoeba Saline before final re-suspension in 1 mL of phosphate buffered saline. Con-centrated cells were placed at -80˚C before DNA extraction and sequencing. Genomic DNA was sequenced using the Illumina MiSeq (Illumina, Inc, San Diego CA 92121, USA).
The gDNA was quantified by a Qubit assay (Life technologies, Carlsbad, CA, USA) to 6.3 ng/μL and dilution was performed requiring 1 ng of DNA as input. The genomic DNA was fragmented and tagged. Limited cycles of PCR amplification completed the tag adapters and introduce dual-index barcodes. After purification on AMPure beads (Life Technologies,
Carlsbad, CA, USA), libraries were then normalized on specific beads according to the Nextera XT protocol (Illumina). Normalized libraries were pooled into a single sequencing library, the MiSeq. The pooled single strand library was loaded onto the reagent cartridge and then onto the instrument along with the flow cell. Automated cluster generation and paired-end sequencing with dual index reads was performed in a single 39-hour, 2x251- base pair run (bp). Total information of 10.5 Gb was obtained from a cluster density of 1,288,000 per mm2 with 86.3% (20,305,000 clusters) of the clusters passing quality control filters. Within this pooled run, the index representation ofT. tenax was determined to 39.43%. The 8,005,980 paired end reads were filtered according to the read qualities.
Multilocus sequence typing. The reads obtained were assembled using the CLC Geno-mics Workbench. Protein sequences were predicted using the Prodigal platform [34] and anal-ysis. A local Blastp on the predicted proteins was performed on NCBI Blast (Basic Local Alignment Search Tool) against non-redundant protein sequences (nr) database using the standard parameters. TheT. tenax NIH4 genome sequence was deposited on the EMBL-EBI website (Bioproject: PRJEB22701 and whole contigs under accession numbers OCT-D01000001-OCTD01004161). Seven single-family household genes were selected ranging in length from 450 to 500 bp (Table 1) based on MLST designed forTrichomonas vaginalis [35]. Fig 1. Schematic protocol forTrichomonas tenax studies.
Single standard PCRs were performed to allow DNA amplification of each selected gene. Primer hybridization was conducted at 59˚C and amplified products were sequenced as described previously [36]. Sequences were corrected and assembled using the ChromasPro software version 1.71 (Technelysium, Australia). Obtained sequences were aligned using MUSCLE tool in the Molecular Evolutionary Genetics Analysis software (MEGA) version 7.0.18 (Pennsylvania State University, United-States) and, finally, phylogenetic trees were gen-erated using the maximum-likelihood (ML) method within FastTree version 2.0 [37]. MLST nucleotide sequences data are available in the EMBL-EBI database under accession numbers LT934459 to LT934497.
Direct molecular detection from clinical samples
qPCR on therpb1 gene was used to confirm the presence of T. tenax directly from clinical samples using specific primers and probe (Fig 1E). Cross amplifications were prevented in silico and primers were tested on the DNA of Trichomonas vaginalis strain G3 (ATCCPRA-98). DNA extraction from our isolates and specimens were performed using the NucleoSpin Tissue kit (Macherey-Nagel, Hoerdt, France). The optimized 20μL Quantitative Real-Time Polymerase Chain Reaction (qPCR) mix contained: master mix (10μL), primers (0.5 μL, 20 nM), probe (0.5μL, 5 nM), water (3.5 μL) and 5 μL of DNA. qPCR temperature cycle was: DNA activation at 50˚C for two minutes, denaturation at 95˚C for five minutes, followed by 40 cycles of 95˚C for one second and 60˚C for 30 seconds for the plate read. qPCR tubes were deposited in a CFX96 Touch thermal cycler (Bio-rad, France). The results were normalized by testing the gene encoding albumin in parallel. The primers used were: Forward: 5’- GCT GTCATCTCTTGTGGGCTG T-3’, Reverse: 5’- AAACTCATGGGAGCTGCTGGTTC-3’ and FAM probe: 6FAM- 5’ CCTGTCATGCCCACACAAATCTCTCC-3’[38]. A ratio of the cycle threshold (Ct) obtained in q-PCR for therpb1 gene and the albumin gene was calculated. (S3 Table).
Statistical analysis
The statistical analysis was performed using “R” software (Version 3.5.1) using a ordinal logis-tic regression with a four-modalities qualitative variable to investigate the correlation between health status and detection methods (culture or PCR) with the existence ofT. tenax.
χ2
tests for sex and smoking status descriptive statistics and Fisher test for the phylogenetic tree analysis usingStatistical Package for the Social Sciences (SPSS Inc, IBM Company). Table 1. Primers used for multilocus sequence typing genes ofTrichomonas tenax.
Genes Forward Reverse Sequence length (bp) No of alleles
Alanyl tRNA synthetase (ALTS) 5’-CCGTCCAGGATGGTGTCTTC-3’ 5’-GTAACATCGAATGGCTGGCAC-3’ 514 5
DNA mismatch repair protein (DMRP) 5’-ATTGGACAATGGAACCAGTCA-3’
5’-TGACCATATTTCGCACCACG-3’ 516 5
Serine hydromethyltransferase (SHMT) 5’-GAGCGATGGAGGACATTTGAC-3’
5’-TTTGGTGAAGATGAGGACCACC-3’
452 4
Mannose-6-phosphate isomerase (M6PI) 5’-AGGTGTTGCAGAGGAGTTGG-3’ 5’-TGCTATTTCGTTTGCAGGAACA-3’
421 7
Glutamine amidotransferase class-I (GAT1)
5’-TCTGTTGCACAAGGTCTCAA-3’ 5’-TTGTGTAGCCGCCGTATTTG-3’ 424 3
Histidyl tRNA synthetase (HIST) 5’-CGTCTCCATCGACACACCAG-3’ 5’-TCCATCTCGTCGAGGACCTT-3’ 658 8
Cysteinyl tRNA synthetase (CYST) 5’-GCCCGACTGTTTACTCGACA-3’ 5’-CGAAGATCGATACCACCGCA-3’ 657 0
Results
Isolation of
Trichomonas sp.
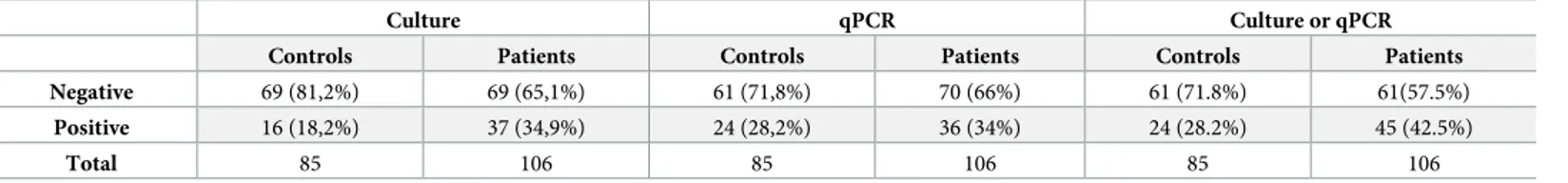
Fifty-three strains ofTrichomonas sp. were obtained from periodontal samples, 37/106 from patients with periodontitis and 16/85 from control patients (p = 0.018) (Table 2,Table 3). Six isolates were lost before conservation. Protists were likely to beT. tenax based on their mor-phology as observed by microscopy. For a definitive identification, we first performed align-ment of availablerpb1 gene using online Clustal Omega [39] with standard parameters for 11 sequences ofTrichomonas spp. and we visualized it using MView online software (http://www. ebi.ac.uk/Tools/msa/mview/). Following the 100% consensus sequence available (S1 Scheme), we designed degenerated primers on conserved regions between the 11 different Trichomona-didae strains. In order to evaluate the intra-species diversity of the rpb1 gene, we amplified and sequenced therpb1 gene from 15 randomly chosen strains isolated from our control and dis-eased patients. Of the 3,000 base pairs obtained, all sequences were 100% identical to the refer-ence strainT. tenax NIH4, except one strain (number 13) which possesses two synonym single nucleotide polymorphisms (SNP). All remaining isolates were identified asT. tenax using our specific primers.
Detection of
T. tenax using quantitative real time PCR
Sixty of the 191 samples were positive forT. tenax, 24/85 controls and 36/106 periodontitis patients (p = 0.089) (Table 2;Table 3). The standardized results using the average ratio of the rpb1 gene on the albumin gene demonstrate the absence of a link between the amount of Trichomonas DNA and the pathological status (p = 0.087, χ2
) (S3 Table).
Combined results
A good correlation was observed between qPCR and culture, as 83% of positive cultures were also positive for qPCR and 88% of negative cultures were also negative for qPCR. By combin-ing both results, 45/106 patients were positive by culture and/or PCR, compared to 24/85 of controls (p = 0.042,χ2
). No association could be found with the patient’s gender and health status (p = 0.710,χ2) or with the detection of T. tenax (p = 0.485, χ2). The carriage ofT. tenax and smoking status are correlated regardless of health status (p = 0.001,χ2). Regarding the periodontitis classification of each patient,T. tenax is significantly more detected in severe periodontitis than in mild or moderate periodontitis (p<0.05).
Table 3. Comparison between frequency of qPCR and culture.
Culture
Negative Positive Total Real-Time PCR Negative 122 (88,4%) 9 (17%) 131
Positive 16 (11,6%) 44 (83%) 60
Total 138 53 191
https://doi.org/10.1371/journal.pone.0213338.t003
Table 2. Frequency ofTrichomonas tenax by qPCR and culture.
Culture qPCR Culture or qPCR
Controls Patients Controls Patients Controls Patients Negative 69 (81,2%) 69 (65,1%) 61 (71,8%) 70 (66%) 61 (71.8%) 61(57.5%)
Positive 16 (18,2%) 37 (34,9%) 24 (28,2%) 36 (34%) 24 (28.2%) 45 (42.5%)
Total 85 106 85 106 85 106
Genome sequencing
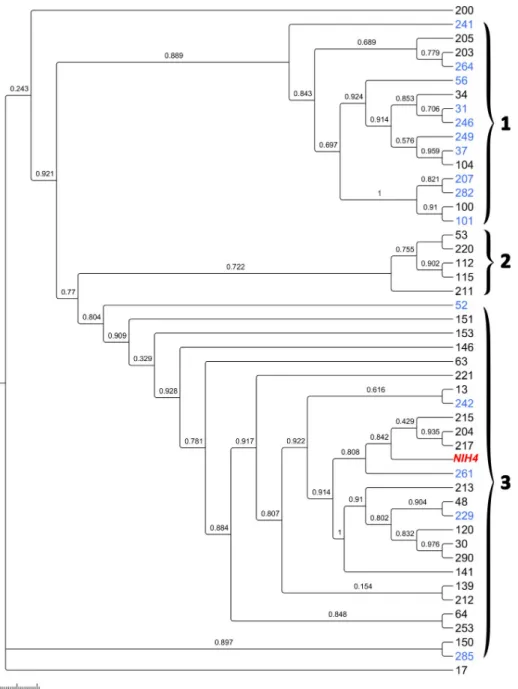
The 46,742,176 base pair (bp) draft genome obtained contains 4,161 scaffolds with a N50 of 13,554 bp with a minimum of 4,002 bp to a maximum of 92,188 bp with a G+C content esti-mated of about 34.6% similar toT. vaginalis G3 (35.5%). The gene prediction determined 34,291 Open Reading Frame (ORF) includes 21,854 proteins that could be annotated and 12,437 ORFans. Of these, 21,193 proteins are in common withT. vaginalis, and only five with Trichomonas gallinae, four with T. tenax (sequences of the same strain already available on the nr database) and one withTritrichomonas foetus. This high number of best hits shared with T. Fig 2.Trichomonas tenax relationship based on multilocus sequence typing phylogenetic analysis. Maximum
likelihood phylogenetic tree ofT. tenax strains. Colors legend: black for patients, blue for control and red for reference
strain.
vaginalis G3 strain is logical, as this species in the only species of Trichomonas spp. for which a draft genome is available. In theT. vaginalis G3 genome, about 250 genes were annotated as ribosomal proteins [40]. InT. tenax, we obtained in the draft 52 ORFs annotated as the 40S ribosome and 68 annotated as the 60S ribosome. We isolated the complete sequence of the rpb1 gene measuring 4,962 nucleotides and, congruently, the alignment with the same strain in the database showed 61% of coverage with 100% identity. The alignment starts at position 229 and finishes at position 3,276 of the complete sequence and makes the 3’end of this gene available.
MLST typing
Four housekeeping genes were extracted based on theT. vaginalis typing system: Alanyl tRNA synthetase (ALTS), DNA mismatch repair protein (DMRP), Serine hydromethyltransferase (SHMT), and Mannose-6-phosphate isomerase (M6PI). For genes for which no homologs of Trichomonas vaginalis could be detected in our T.tenax draft genome, we chose three genes because of their functional similarity to complete the system: Glutamine amidotransferase class-I (GAT1), Histidyl tRNA synthetase (HIST), Cysteinyl tRNA synthetase (CYST) (Table 1). The 47 strains and the reference NIH4 strain are clustered into three major groups: strains obtained in the control group are significantly predominant in group 1 and strains iso-lated in periodontitis patients are significantly predominant in group 3 (p <0.05, Fisher). Only periodontal patient strains constitute the cluster 2. (Fig 2). Phylogenetic tree analysis, based on each housekeeping gene, showed similar topologies. Indeed, six of seven genes also present three major clusters, namely the DMRP, SHMT, ALTS, SHMT, M6PI and GAT1 gene (S1–S7
Figs). Concerning the tree based on the CYST gene, all sequences are identical between con-trols and patients, highlighting the conservation and lack of variability in this portion of the gene. All strain sequences obtained for each gene portion were compared with the sequences of the reference strainT. tenax NIH4 to investigate the presence of a single nucleotide poly-morphism (SNP) (S2 Table). Four isolates had the same genotype, while the others 43 isolates each had a specific genotype.
Discussion
In this study, by combining a polyphasic approach that associates culture and qPCR, we found a correlation between periodontitis and the presence ofT. tenax (p< 0.05). Although T. tenax is more frequently detected by qPCR in patients than in controls, the difference is not signifi-cant (p = 0.435). A signifisignifi-cant difference was observed using culture only (p = 0.015). By com-bining the culture and PCR results to neutralize the effect of false negative of each technique and evaluate the real prevalence ofT. tenax, the difference is significant (p = 0.042). The proba-bility of false positive/negative frequency is reduced due to the good correlation observed between both techniques: 83% of positive cultures also positive for qPCR and 88% of negative cultures also negative for q-PCR techniques. We believe that this good correlation indicates that the culture and handling protocols, including the transport medium specifically developed for anaerobic microorganisms, were highly efficient. However, as commonly observed in clini-cal microbiology, the higher sensitivity of the PCR suggests that someT. tenax did not grew in culture. The reasons are unknown but usually because microorganisms are dead at time of inoculation due to delayed inoculation between sampling and culture, quality of the operator or quality of the batch of transport or culture media. The 11.6% of the positive samples in cul-ture not identified by real-time PCR shows that false positive occur also with molecular ampli-fication, usually as a consequence of inhibitors.
Furthermore, positive culture in controls underlines the difficulty of having a true negative control group. We first suspected that a higher rate of positive culture in periodontitis patients could have been the result of a higher concentration of protist in these samples. But the absence of such differences observed using quantitative PCR disproves this hypothesizes. Indeed, no difference in the amount ofT. tenax DNA in controls and patients is demonstrated after normalization of the results by the albumin gene. A recent review reports higher preva-lence ofT. tenax occurrence in gum diseased (gingivitis and periodontitis) in most studies present in the literature and explains that the heterogeneity of the prevalence observed may be due to the different methods used to detect the protist (majority of microscopic observation, insufficient use of molecular biology) and the studied population diversity [21]. The same lim-its may be pointed out in another study reporting the absence of trichomonas in healthy sites in periodontitis patients [27]. Theses discrepancies, as compared to our study, could be related with false positive of our PCR procedure. However, the high level of positive culture herein is not in agreement with this hypothesis and the detection of trichomonads from healthy sites in controls with no periodontal disease indicated thatT. tenax carriage remains common in the oral cavity.
A correlation could also be established between the severity of periodontitis and the pres-ence of protists (p<0.05).T. tenax is found in severe periodontitis differing from other peri-odontitis, by the depth of the pocket as well as the loss of attachment. The environment of severe periodontitis would therefore be more favorable to the development of the protist and other bacteria species leading to serious lesions and inflammatory responses.
We now suspect that the specific periodontal microbiota associated with periodontitis could promote the growth ofT. tenax. Indeed, when inoculated in the culture medium, the sample also contains bacterial microbiota of dental pockets. Indeed, periodontitis is suspected to be due to an inflammatory response to microorganisms [1,8]. Finally, no single microorgan-ism is implicated but rather a combination of microorganmicroorgan-isms act synergistically [3,17,20]. Investigating the difference in microbiota between healthy individuals and patients with mild and moderate periodontitis compared to severe periodontitis would highlight the combined role of protist and oral microbiome.
Several studies have demonstrated the efficacy of PCR in detectingT. tenax by employing rRNA sequencing [41]. Likewise, oral cavity metagenomics studies have identified significant T. tenax rRNA intergenic spacers [42]. Nonetheless, these systems come up against identifica-tion limits. Designing primers based on therpb1 gene according to Malik et al. [33], allowed us to create a highly specific and sensitive system: this gene is both discriminatory between dif-ferent species and genotypes and is highly conserved within the same species [43,44]. All iso-lated strains ofTrichomonas sp. were identified as T. tenax, once again demonstrating the effectiveness of this primer system and the extreme conservation ofT. tenax based on the rpb1 gene.
Finally, the MLST system made it possible to investigate the clonal relationship between the protist and periodontitis based on the system created for the closely studied neighboring spe-cies,T. vaginalis [35]. The data obtained reveal the existence of three clusters groupingT. tenax, suggesting genetically diverse strains affecting the periodontium tissue. A significant association could be observed between clustering and the occurrence of periodontitis. The pos-sibility of finding new virulence factors in common between strains in future work could con-firm the virulence of diseased strains related to the severity of the periodontal disease.
Recently, a physiopathology mechanism ofT. tenax has been described in the periodontitis disease:T. tenax induced an effect against the human macrophage and deregulates the proin-flammatory cytokines [45]. Furthermore,in vitro studies showed that T. tenax had cytotoxic effects on mammalian cells [46]. An animal model reproducing periodontitis suggested in
these recent studies could be used to explore the potential pathophysiological role ofT. tenax [21,47].
Conclusion
A high prevalence ofT. tenax in both controls and patients is detected using genomic-depen-dent and culture-based methods of detection.T. tenax was more frequently associated with severe periodontitis. Three clusters of strains were highlighted by the MLST genotyping sys-tem, two were significantly associated with periodontitis.T. tenax appears to be associated with the onset or/and evolution of periodontal diseases. However, although these differences are statistically significant, it is impossible to determine whether they are a cause or a conse-quence of the disease.
Supporting information
S1 Scheme. Quantitative Real-Time PCR (q-RT PCR) specific forTrichomonas tenax.
(DOCX)
S1 Table. The table below presents the classification of each patient. The numbers corre-spond to a patient and preserve anonymity. The discontinuity of the numbering is linked to the change of operator in the laboratory. In black: patients with periodontitis.—In bold: the patient controls.
(DOCX)
S2 Table. Sequence types ofTrichomonas tenax based on single-nucleotide polymorphism
against theTrichomonas tenax reference strain. In bold: the number of sequences types.
(DOCX)
S3 Table. The table below presents the cycle threshold obtained in q-PCR for theRPB1gene
and the albumin gene for the standardization. In black: periodontitis patients. In bold: con-trols.
(DOCX)
S1 Fig.Trichomonas tenax phylogenetic analysis based on Alanyl tRNA synthetase (ALTS)
gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S2 Fig.Trichomonas tenax phylogenetic analysis based on DNA mismatch repair protein
(DMRP) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S3 Fig.Trichomonas tenax phylogenetic analysis based on Serine hydromethyltransferase
(SHMT) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S4 Fig.Trichomonas tenax phylogenetic analysis based on Mannose -6-phosphate
isomer-ase (M6PI) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S5 Fig.Trichomonas tenax phylogenetic analysis based on Glutamine amidotransferase
class-I (GAT1) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S6 Fig.Trichomonas tenax phylogenetic analysis based on Histidyl tRNA synthetase (HIST) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
S7 Fig.Trichomonas tenax phylogenetic analysis based on Cysteinyl tRNA synthetase
(CYST) gene. In black: patients with periodontitis. In blue: the patient controls. (TIFF)
Acknowledgments
The authors extend their thanks to Emeline Baptiste for the genome and sequences submis-sions and Caroline Michelle for sequencing.
Author Contributions
Conceptualization: Gerard Aboudharam, Bernard La Scola.
Formal analysis: Sarah Benabdelkader, Julien Andreani, Herve Chaudet, Bernard La Scola. Investigation: Sarah Benabdelkader, Julien Andreani, Alexis Gillet, Elodie Terrer, Marion
Pignoly.
Methodology: Herve Chaudet.
Project administration: Bernard La Scola. Supervision: Gerard Aboudharam. Validation: Bernard La Scola.
Writing – original draft: Sarah Benabdelkader, Julien Andreani. Writing – review & editing: Gerard Aboudharam, Bernard La Scola.
References
1. Pihlstrom BL, Michalowicz BS, Johnson NW. Periodontal diseases. Lancet (London, England). 2005; 366: 1809–20.https://doi.org/10.1016/S0140-6736(05)67728-8
2. Tonetti MS, Greenwell H, Kornman KS. Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J Clin Periodontol. 2018; 45: S149–S161.https://doi.org/10. 1111/jcpe.12945PMID:29926495
3. Socransky SS, Haffajee a D, Cugini M a, Smith C, Kent RL. Microbial complexes in subgingival plaque. J Clin Periodontol. 1998; 25: 134–144.https://doi.org/10.1111/j.1600-051X.1998.tb02419.xPMID: 9495612
4. Rosier BT, De Jager M, Zaura E, Krom BP. Historical and contemporary hypotheses on the develop-ment of oral diseases: are we there yet? Front Cell Infect Microbiol. 2014;https://doi.org/10.3389/fcimb. 2014.00092PMID:25077073
5. Assuma R, Oates T, Cochran D, Amar S, Graves DT. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J Immunol. 1998; 160: 403–9. PMID:9551997 6. Nociti FH, Casati MZ, Duarte PM. Current perspective of the impact of smoking on the progression and
treatment of periodontitis. Periodontol 2000. 2015; 67: 187–210.https://doi.org/10.1111/prd.12063 PMID:25494601
7. Lalla E, Papapanou PN. Diabetes mellitus and periodontitis: a tale of two common interrelated diseases. Nat Rev Endocrinol. 2011; 7: 738–48.https://doi.org/10.1038/nrendo.2011.106PMID:21709707 8. Kinane DF, Stathopoulou PG, Papapanou PN. Periodontal diseases. Nat Rev Dis Prim. Macmillan
Pub-lishers Limited; 2017; 3: 17038.https://doi.org/10.1038/nrdp.2017.38PMID:28805207
9. Ge X, Rodriguez R, Trinh M, Gunsolley J, Xu P. Oral microbiome of deep and shallow dental pockets in chronic periodontitis. PLoS One. Public Library of Science; 2013; 8: e65520.https://doi.org/10.1371/ journal.pone.0065520PMID:23762384
10. Wang J, Qi J, Zhao H, He S, Zhang Y, Wei S, et al. Metagenomic sequencing reveals microbiota and its functional potential associated with periodontal disease. Sci Rep. Nature Publishing Group; 2013; 3: 1843.https://doi.org/10.1038/srep01843PMID:23673380
11. Griffen AL, Beall CJ, Campbell JH, Firestone ND, Kumar PS, Yang ZK, et al. Distinct and complex bac-terial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J. Nature Pub-lishing Group; 2012; 6: 1176–85.https://doi.org/10.1038/ismej.2011.191PMID:22170420
12. Galimanas V, Hall MW, Singh N, Lynch MD, Goldberg M, Tenenbaum H, et al. Bacterial community composition of chronic periodontitis and novel oral sampling sites for detecting disease indicators. Microbiome. 2014; 2.https://doi.org/10.1186/2049-2618-2-32PMID:25225610
13. Kumar PS, Griffen AL, Barton JA, Paster BJ, Moeschberger ML, Leys EJ. New bacterial species associ-ated with chronic periodontitis. J Dent Res. 2003; 82: 338–344.https://doi.org/10.1177/
154405910308200503PMID:12709498
14. Bik EM, Long CD, Armitage GC, Loomer P, Emmerson J, Mongodin EF, et al. Bacterial diversity in the oral cavity of ten healthy individuals. Isme J. 2010; 4: 962–974.https://doi.org/10.1038/ismej.2010.30 PMID:20336157
15. Ly M, Abeles SR, Boehm TK, Robles-Sikisaka R, Naidu M, Santiago-Rodriguez T, et al. Altered Oral Viral Ecology in Association with Periodontal Disease. MBio. 2014; 5: e01133-14–e01133-14.https:// doi.org/10.1128/mBio.01133-14PMID:24846382
16. Duran-Pinedo AE, Chen T, Teles R, Starr JR, Wang X, Krishnan K, et al. Community-wide transcrip-tome of the oral microbiome in subjects with and without periodontitis. ISME J. Nature Publishing Group; 2014; 8: 1659–72.https://doi.org/10.1038/ismej.2014.23PMID:24599074
17. Cappuyns I, Gugerli P, Mombelli A. Viruses in periodontal disease—a review. Oral Dis. Munksgaard International Publishers; 2005; 11: 219–229.https://doi.org/10.1111/j.1601-0825.2005.01123.xPMID: 15984953
18. Hersh SM. Pulmonary trichomoniasis and Trichomonas tenax. J Med Microbiol. 1985; 20: 1–10.https:// doi.org/10.1099/00222615-20-1-1PMID:3894667
19. Lyons T, Scholten T, Palmer JC, Stanfield E. Oral amoebiasis: the role of Entamoeba gingivalis in peri-odontal disease. Quintessence Int Dent Dig. 1983; 14: 1245–1248. PMID:6585855
20. Bonner M, Amard V, Bar-Pinatel C, Charpentier F, Chatard J-M, Desmuyck Y, et al. Detection of the amoeba Entamoeba gingivalis in periodontal pockets. Parasite. 2014; 21: 30.https://doi.org/10.1051/ parasite/2014029PMID:24983705
21. Marty M, Lemaitre M, Ke´moun P, Morrier J, Monsarrat P. Trichomonas tenax and periodontal diseases: a concise review. 2017;https://doi.org/10.1017/S0031182017000701PMID:28583214
22. Canabarro A, Valle C, Farias MR, Santos FB, Lazera M, Wanke B. Association of subgingival coloniza-tion of Candida albicans and other yeasts with severity of chronic periodontitis. J Periodontal Res. 2013; 48: 428–32.https://doi.org/10.1111/jre.12022PMID:23137301
23. Cepicka I, Hampl V, Kulda J. Critical Taxonomic Revision of Parabasalids with Description of one New Genus and three New Species. Protist. 2010; 161: 400–433.https://doi.org/10.1016/j.protis.2009.11. 005PMID:20093080
24. Brook B, Schuster FL. Oral Protozoa: Survey, Isolation, and Ultrastructure of Trichomonas tenax from Clinical source. Trans Am Microsc Soc. 1984; 103: 376–382.
25. Maritz JM, Land KM, Carlton JM, Hirt RP. What is the importance of zoonotic trichomonads for human health? Trends Parasitol. Elsevier Ltd; 2014; 30: 333–341.https://doi.org/10.1016/j.pt.2014.05.005 PMID:24951156
26. Ghabanchi J, Zibaei M, Afkar MD, Sarbazie AH. Prevalence of oral Entamoeba gingivalis and Tricho-monas tenax in patients with periodontal disease and healthy population in Shiraz, southern Iran. Indian J Dent Res. 2010; 21: 89–91.https://doi.org/10.4103/0970-9290.62821PMID:20427914
27. Bisson C, Lec P-H, Blique M, Thilly N, Machouart M. Presence of trichomonads in subgingival biofilm of patients with periodontitis: preliminary results. Parasitol Res. 2018; https://doi.org/10.1007/s00436-018-6077-2PMID:30215136
28. Duboucher C, Mogenet M, Pe´rie´ G. Salivary trichomoniasis. A case report of infestation of a submaxil-lary gland by Trichomonas tenax. Arch Pathol Lab Med. 1995; 119: 277–9. PMID:7887784
29. Duboucher C, Farto-Bensasson F, Che´ ron M, Peltier JY, Beaufils F, Pe´rie´ G. Lymph node infection by Trichomonas tenax: report of a case with co-infection by Mycobacterium tuberculosis. Hum Pathol. 2000; 31: 1317–21.https://doi.org/10.1053/hupa.2000.18502PMID:11070125
30. Lewis KL, Doherty DE, Ribes J, Seabolt JP, Bensadoun ES. Empyema caused by trichomonas. Chest. 2003; 123: 291–2. PMID:12527635
31. Mehr AK, Zarandi A, Anush K. Prevalence of Oral Trichomonas tenax in Periodontal Lesions of Down Syndrome in Tabriz, Iran. J Clin Diagn Res. 2015; 9: ZC88–90.https://doi.org/10.7860/JCDR/2015/ 14725.6238PMID:26393213
32. Eke PI, Page RC, Wei L, Thornton-Evans G, Genco RJ. Update of the Case Definitions for Population-Based Surveillance of Periodontitis. J Periodontol. 2012; 83: 1449–1454.https://doi.org/10.1902/jop. 2012.110664PMID:22420873
33. Malik SB, Brochu CD, Bilic I, Yuan J, Hess M, Logsdon JM, et al. Phylogeny of parasitic parabasalia and free-living relatives inferred from conventional markers vs. Rpb1, a single-copy gene. PLoS One. 2011; 6.https://doi.org/10.1371/journal.pone.0020774PMID:21695260
34. Hyatt D, Chen G-L, Locascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recogni-tion and translarecogni-tion initiarecogni-tion site identificarecogni-tion. BMC Bioinformatics. 2010; 11: 119.https://doi.org/10. 1186/1471-2105-11-119PMID:20211023
35. Cornelius DC, Robinson DA, Muzny CA, Mena LA, Aanensen DM, Lushbaugh WB, et al. Genetic char-acterization of Trichomonas vaginalis isolates by use of multilocus sequence typing. J Clin Microbiol. 2012; 50: 3293–3300.https://doi.org/10.1128/JCM.00643-12PMID:22855512
36. Benamar S, Cassir N, Merhej V, Jardot P, Robert C, Raoult D, et al. Multi-spacer typing as an effective method to distinguish the clonal lineage of Clostridium butyricum strains isolated from stool samples during a series of necrotizing enterocolitis cases. J Hosp Infect. 2017; 95: 300–305.https://doi.org/10. 1016/j.jhin.2016.10.026PMID:27988045
37. Price MN, Dehal PS, Arkin AP. Fasttree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol Biol Evol. 2009; 26: 1641–1650.https://doi.org/10.1093/molbev/msp077 PMID:19377059
38. Carcopino X, Henry M, Benmoura D, Fallabregues AS, Richet H, Boubli L, et al. Determination of HPV type 16 and 18 viral load in cervical smears of women referred to colposcopy. J Med Virol. 2006; 78: 1131–40.https://doi.org/10.1002/jmv.20673PMID:16789021
39. Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol. 2011; 7: 539.https://doi.org/ 10.1038/msb.2011.75PMID:21988835
40. Carlton JM, Hirt RP, Silva JC, Delcher AL, Schatz M, Zhao Q, et al. Draft Genome Sequence of the Sex-ually Transmitted Pathogen Trichomonas vaginalis. Alias J Smith Terry R Utterback Kazutoyo Osoe-gawa Pieter J Jong John M Logsdon Jr Sci. 2007; 11: 207–212.
41. Mallat H, Podglajen I, Lavarde V, Mainardi J, Frappier J, Cornet M. Molecular Characterization of Trich-omonas tenax Causing Pulmonary Infection. 2004; 42: 3886–3888.https://doi.org/10.1128/JCM.42.8. 3886
42. Belda-Ferre P, Alcaraz LD, Cabrera-Rubio R, Romero H, Simo´n-Soro A, Pignatelli M, et al. The oral metagenome in health and disease. ISME J. Nature Publishing Group; 2012; 6: 46–56.https://doi.org/ 10.1038/ismej.2011.85PMID:21716308
43. Bilic I, Jaskulska B, Souillard R, Liebhart D, Hess M. Multi-Locus Typing of Histomonas meleagridis Iso-lates Demonstrates the Existence of Two Different Genotypes. Leung FCC, editor. PLoS One. Public Library of Science; 2014; 9: e92438.https://doi.org/10.1371/journal.pone.0092438PMID:24658534 44. Hussain I, Jaskulska B, Hess M, Bilic I. Detection and quantification of Histomonas meleagridis by
real-time PCR targeting single copy genes. Vet Parasitol. 2015; 212: 382–388.https://doi.org/10.1016/j. vetpar.2015.08.011PMID:26319200
45. Govro EJ, Stuart MK. Cytokine response of human THP-1 macrophages to Trichomonas tenax. Exp Parasitol. 2016; 169: 77–80.https://doi.org/10.1016/j.exppara.2016.07.011PMID:27497807
46. Ribeiro LC, Santos C, Benchimol M. Is Trichomonas tenax a Parasite or a Commensal? Protist. Elsevier GmbH.; 2015; 166: 196–210.https://doi.org/10.1016/j.protis.2015.02.002PMID:25835639
47. Lemaitre M, Monsarrat P, Blasco-Baque V, Loubières P, Burcelin R, Casteilla L, et al. Periodontal Tis-sue Regeneration Using Syngeneic Adipose-Derived Stromal Cells in a Mouse Model. Stem Cells Transl Med. 2017; 6: 656–665.https://doi.org/10.5966/sctm.2016-0028PMID:28191762