Folliculin regulates mTORC1/2 and WNT pathways in early human pluripotency
14
0
0
Texte intégral
Figure
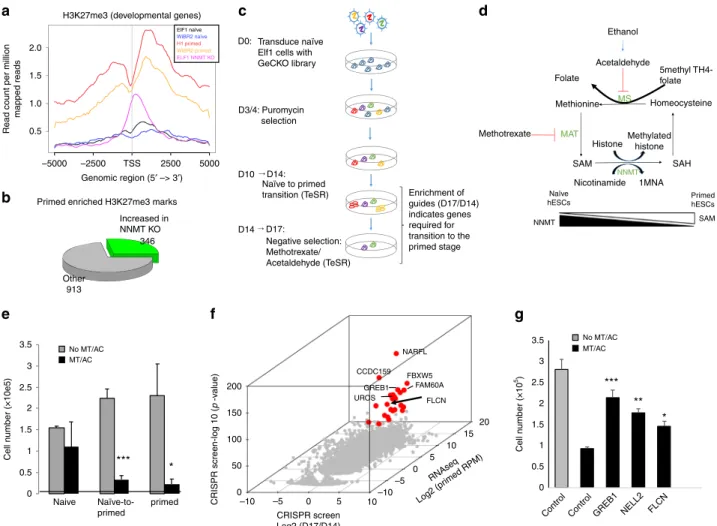
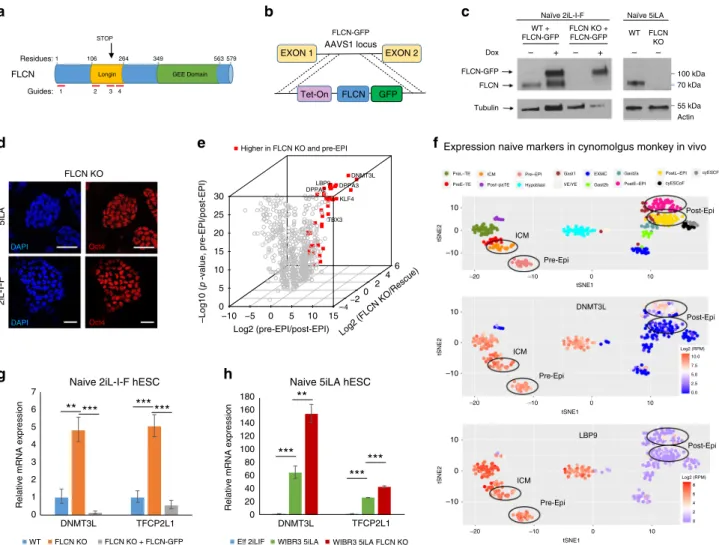
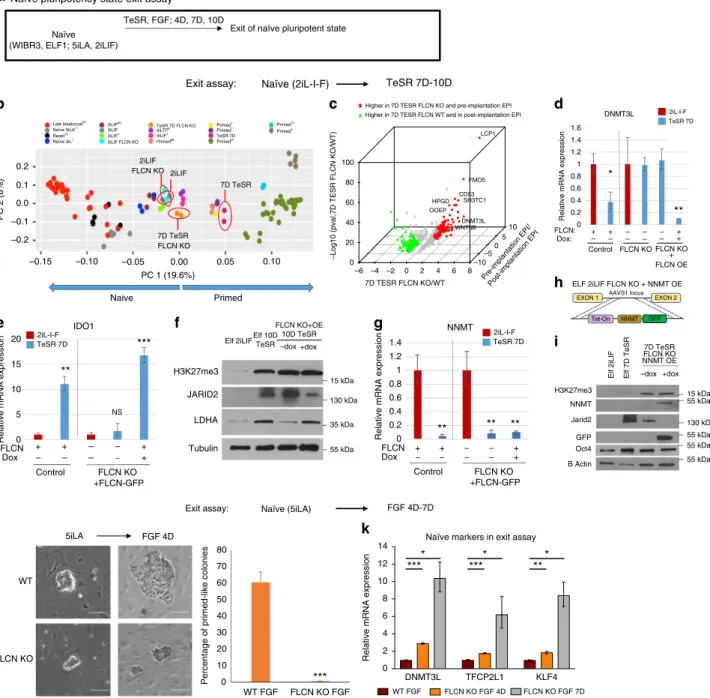
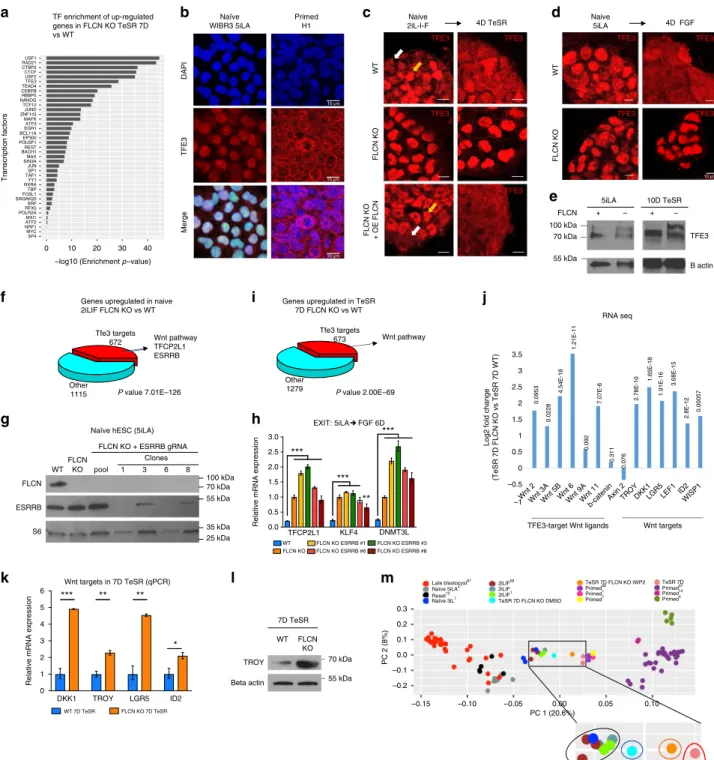
Documents relatifs