Deep Chlorophyll Maxima in the Global Ocean: Occurrences, Drivers and Characteristics
Texte intégral
Figure
![Figure 1. Map of the BGC-Argo profiles where the [Chla] and the b bp are measured. BGC, Biogeochemical.−50050−150−100 −50 0 50 100 150Longitude (°E)Latitude (°N)](https://thumb-eu.123doks.com/thumbv2/123doknet/14793478.602524/5.918.67.427.865.1063/figure-argo-profiles-chla-measured-biogeochemical-longitude-latitude.webp)
![Figure 2. Example of vertical profiles of [Chla] (green) and b bp (blue) classified as (a), (b) NO, (c) DAM, and (d) DBM](https://thumb-eu.123doks.com/thumbv2/123doknet/14793478.602524/6.918.286.841.134.440/figure-example-vertical-profiles-chla-green-blue-classified.webp)




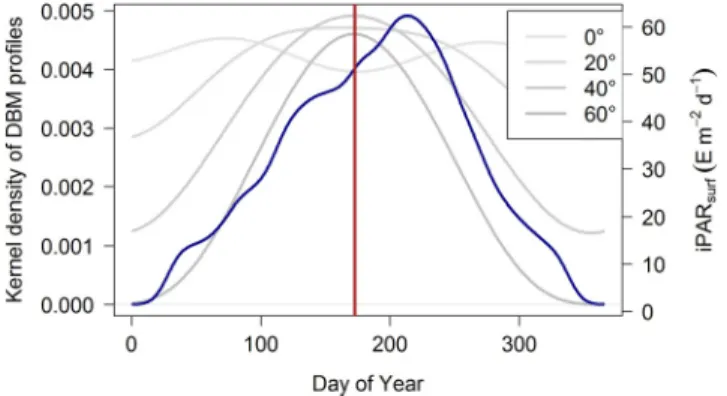
![Figure 13. Quartile diagrams of (a) MLD, (b) [Chla] sat , (c) mean daily PAR in the mixed layer ( iPAR ML ), and (d) mean nitrate concentration in the ML ( Nitrate ML ) per Northern Hemisphere-phased month (regardless of the year) for DAZ profiles with](https://thumb-eu.123doks.com/thumbv2/123doknet/14793478.602524/20.918.285.842.139.811/quartile-diagrams-concentration-nitrate-northern-hemisphere-regardless-profiles.webp)
Documents relatifs
tial cross section is noticeably reduced by increasing column density. If a high col- umn density reference spectrum from the lab were used for DOAS retrieval of low column
The serious changes in ELF magnetic field intensity were detected on 20 Septem- ber 1999, in such a way that the ELF noise level is found to increase by more than 5 dB from the
While devices can be engineered to control the drug release rate, the design of implantable strategies should be based on an understanding that bioavailability of a drug released
Une étude a prouvé qu’une éducation thérapeutique axée sur les connaissances de la pathologie ainsi que sur les autosoins chez les patients souffrant d’insuffisance cardiaque
Copier un texte sans
magnesium silicate phases and water storage capacity in the transition zone and lower 487. mantle, Phys. Nestola, McNeill J., Nasdala L., Hutchison M. The Macusani glasses
physical and biogeochemical processes led to an increase of the nutrient stock in the upper layer 621. (Fig. 7), thickening the ML to 50-60 m depth on average, close to the depth
Within this general objective, three main topics are addressed: (1) we investigate to what degree the increase of recurrent surface [Chl a] during winter (as seen from satellite
