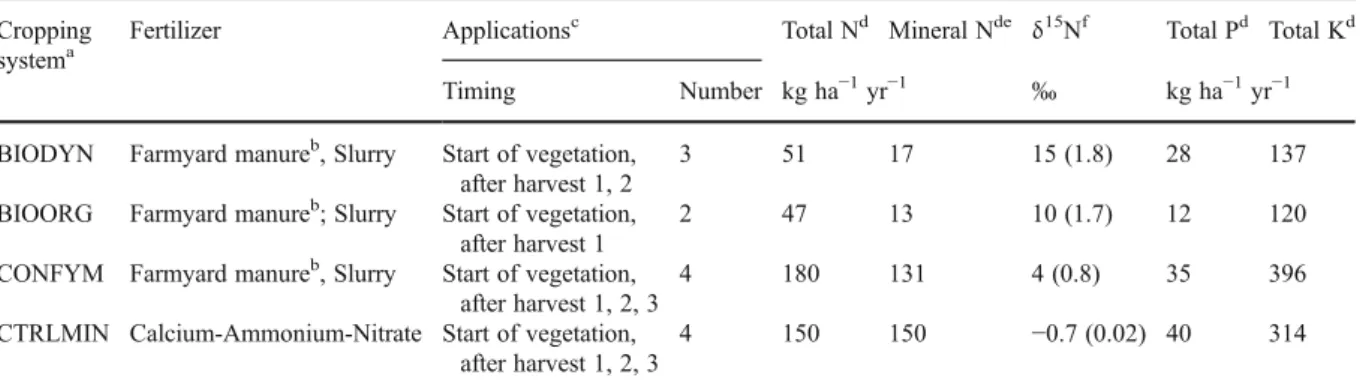
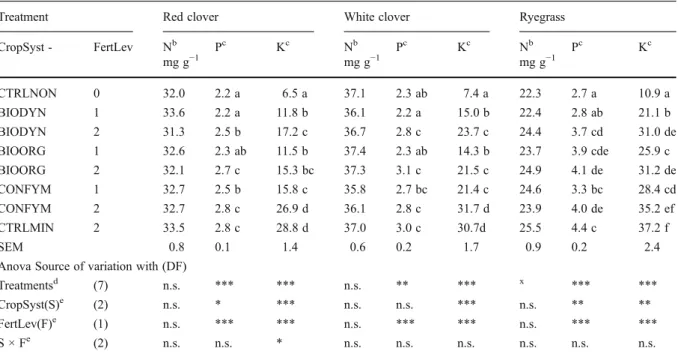
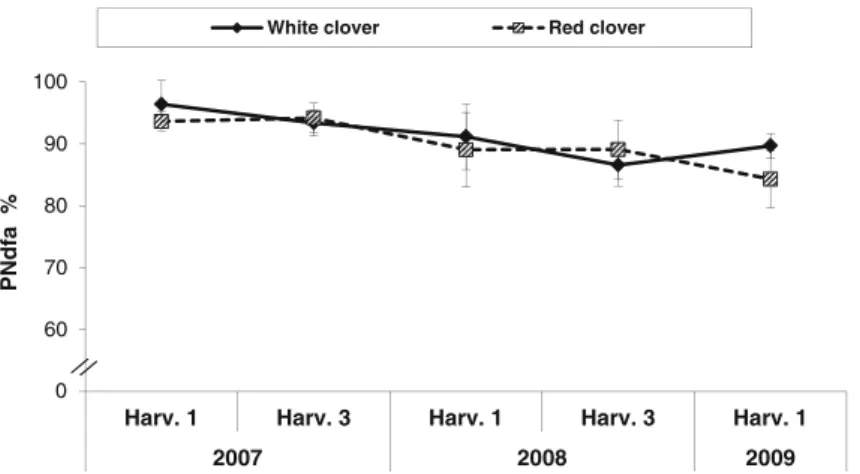
Nitrogen fixation and transfer in grass-clover leys under organic and conventional cropping systems
Texte intégral
Figure

Documents relatifs
The degree of adoption of management techniques involving the increase of organic matter inputs to the soil exerted a profound influence on the total global warming potential
Density (number m −2 ) of the total community and the three ecological groups in the three cropping systems (“direct seeding living mulch-based cropping system”, named living
This paper presents the prediction of the Moorepark Grass Growth model (MGGm) in terms of grass growth and nitrogen content of the grass in a grazing context with different
This study involved the creation of a dynamic and mechanistic grass growth model (the Moorepark Grass Growth Model; hereafter referred to as MGGM) by adapting an existing grass
The adapted version of the Jouven Model can be used for grass growth simulation; however, further changes to the model could be made around the functions of the
In conclusion, short-term CA practices in the three cropping systems increased the storage of STN, SOC and labile SOC pool and enhanced soil enzyme activities in the surface soils
parameters under different systems present in savannah grasslands, pine forest, rice after soil 8.. ploughing, and fodder species (Brachiaria ruziziensis, Stylosanthes
In this study, the soil management techniques differ essentially by soil tillage and soil cover: (1) traditional seeding on tilled soil (TS), (2) seeding on non tilled soil (NS),
