Alleviating Pain Hypersensitivity through Activation of Type 4 Metabotropic Glutamate Receptor
Texte intégral
Figure
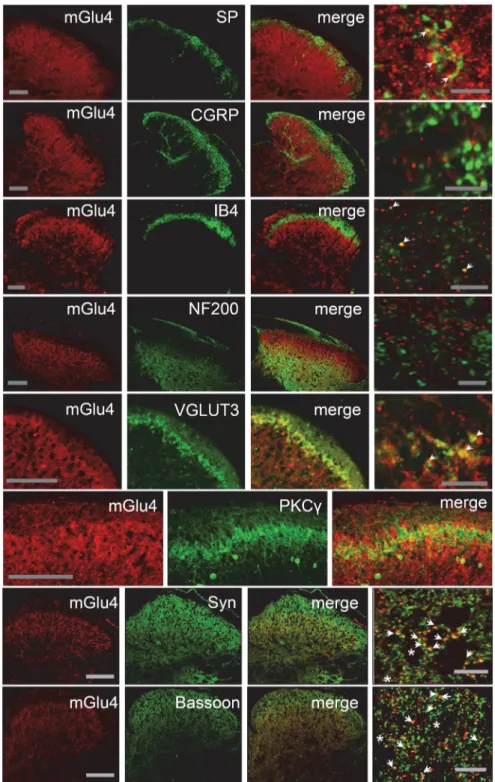
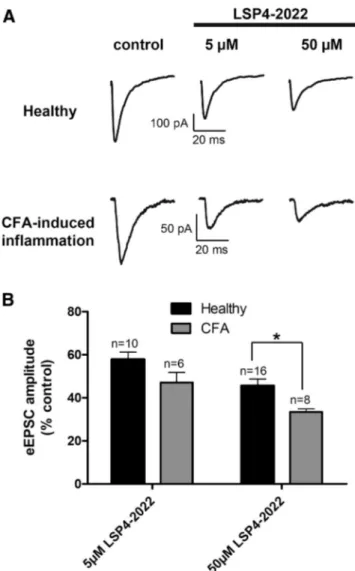
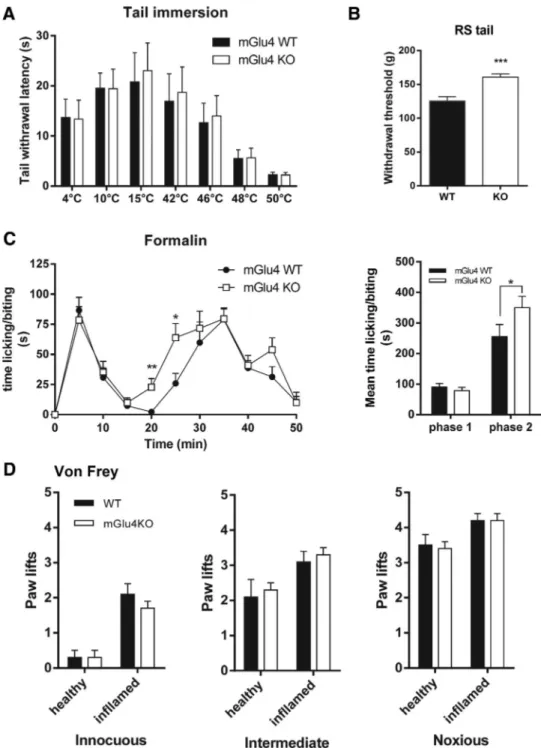
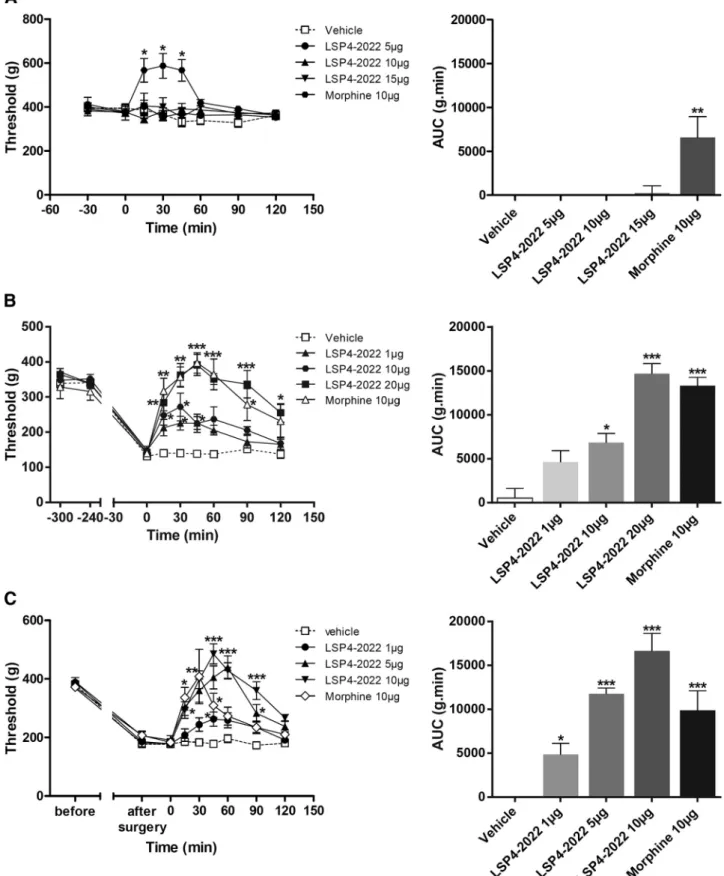
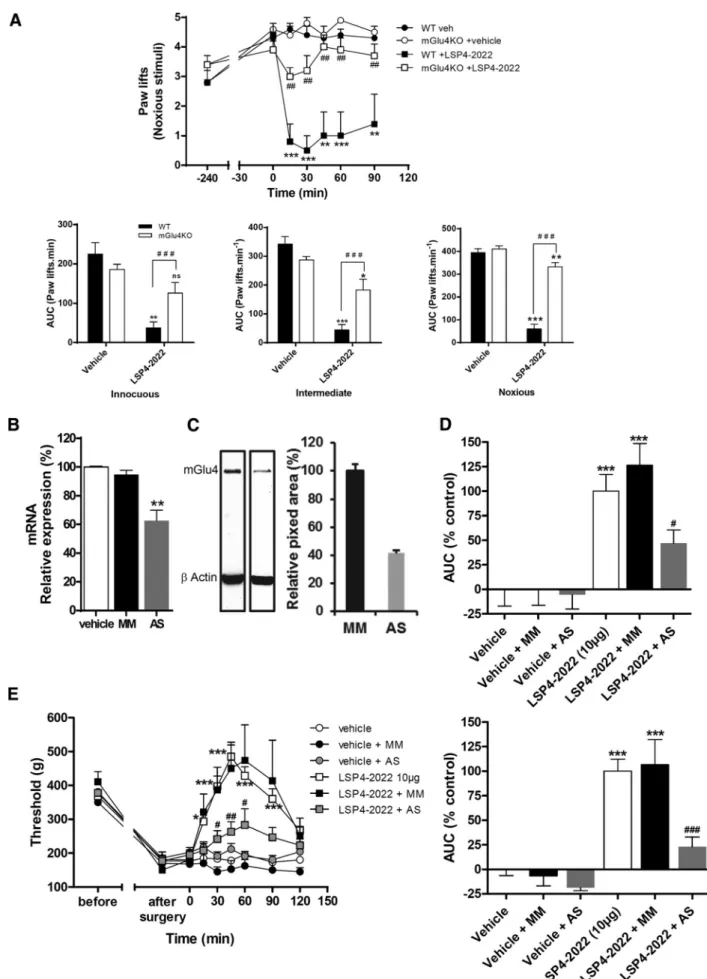
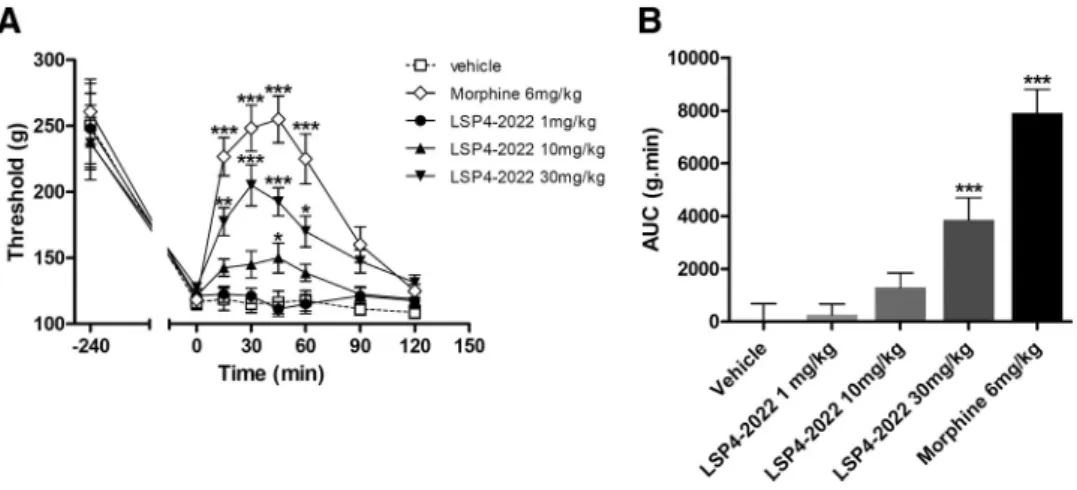
Documents relatifs
Blocking mGluRs prevented the increase in the PPF ratio induced by DL-TBOA in slices from aged rats, indicating that a facilitation of presynaptic mGluR activation can be achieved
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB..
These findings suggest that the procedure can be effective in the rapid treatment of disruptive, aggressive behavior in junior high level special class situations..
Chaque fois qu’il va à la pêche, il revient avec de très gros poissons qu’il partage avec ses voisins.. Ses qualités ont fait de lui une
Proportionnellement, les laboureurs et manouvriers ensemble sont plus nombreux à Montavon (86.5%) que dans le reste de la Vallée (61.5%), qui compte des métiers absents de
Cryptocurrency champions and representatives of the financial services industry do agree with each other that the blockchain technology is a disruptive innovation
In addition, a pilot study with 12 AA obese breast cancer survivors showed that this published dietitian-led approach resulted in a mean loss of 9.8% of baseline
This effect remained significant (p < 0.001) two days after the second injection (Fig. We then investigated whether the receptor was expressed in the DRG and/or
