Calretinin-immunoreactivity during postnatal development of the rat isocortex: a qualitative and quantitative study
13
0
0
Texte intégral
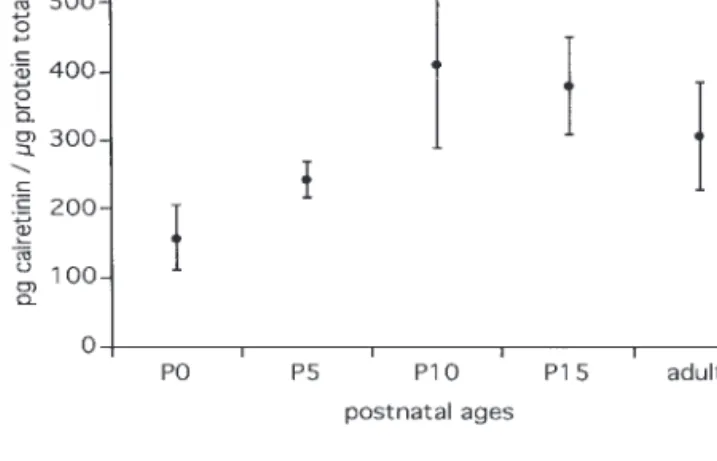
Figure
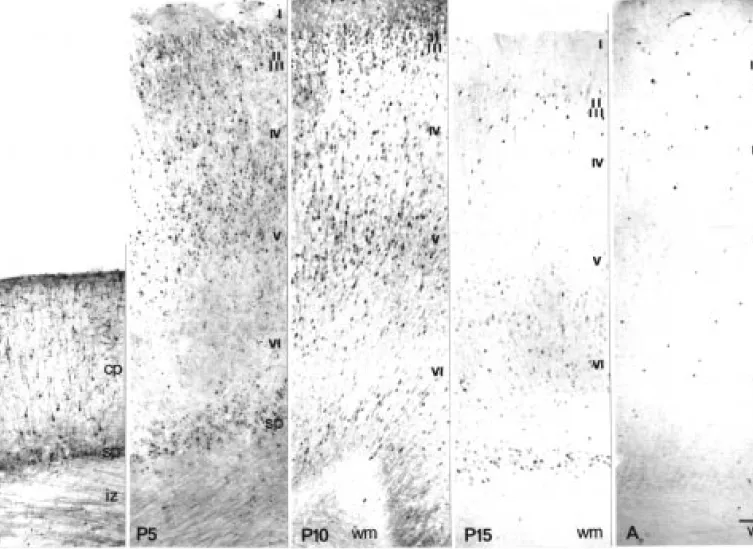
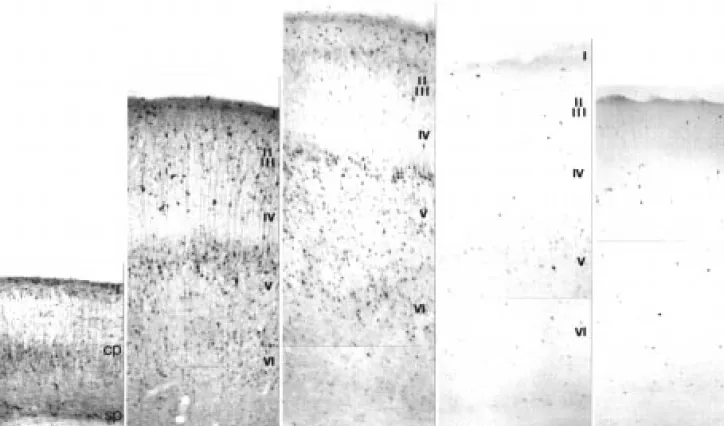

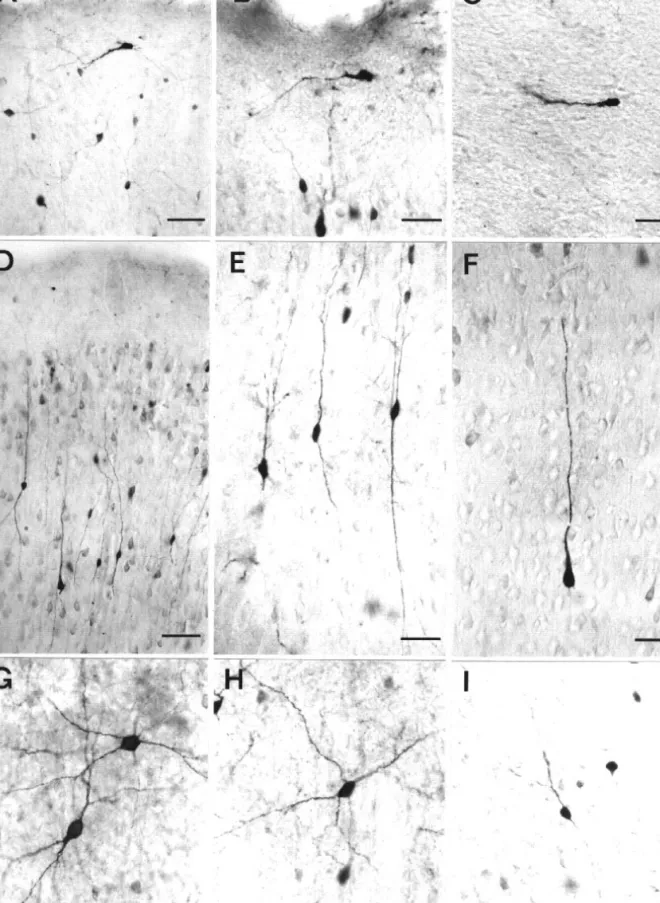
+3

Documents relatifs