Polyphemus: a Palaeolithic Tale?
Texte intégral
Figure


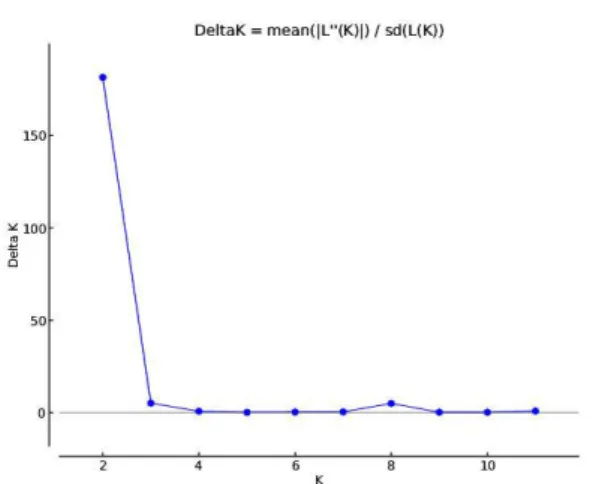


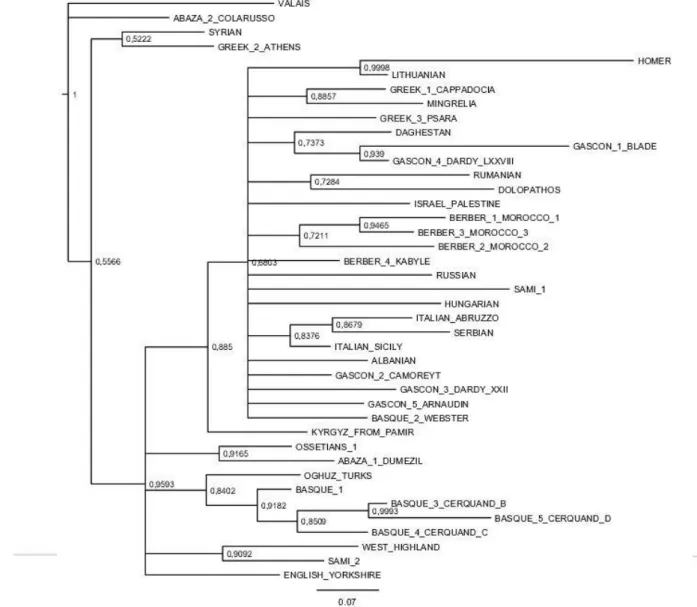
Documents relatifs
triggered by September 2001 (one hesitates to imagine the state of the American body politic after a major terrorist attack every year for nearly a decade) contains
“[m]ore so than any other American author of the twentieth century, Pynchon documents in fiction the tectonic movements of American history, to the extent that
[ 1 ] High-resolution studies of variations in the elemental and stable carbon- and nitrogen-isotope composition of organic matter in cores from Lakes Malawi, Tanganyika, and
KEYWORDS: global legal standards, economic crisis, capitalism, global economic policy, legal theory, finance, financial regulation... This article was originally published under
The pri- mary and secondary wave crests at CWG 5 to CWG 7 have a small temporal offset of Dt = 0.1 s between the peaks compared between both test conditions (Fig. 12 e–g), yet
A genuine economic and monetary union requires establishing both a “fiscal centre” with fiscal capacity to meet the challenge of solidaristic reconstruction and transformation of
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
In summary, the reckless initiative of describing new fossil turtle species based on fragmentary shell material lacking heterobathmy of characters with its sister group, (as
