Reproductive Modes in Onion Thrips (Thysanoptera: Thripidae) Populations from New York Onion Fields
8
0
0
Texte intégral
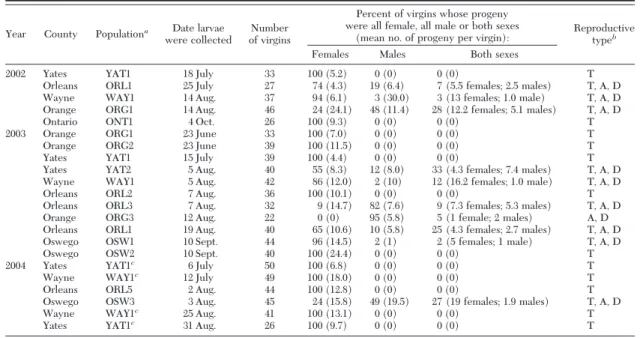
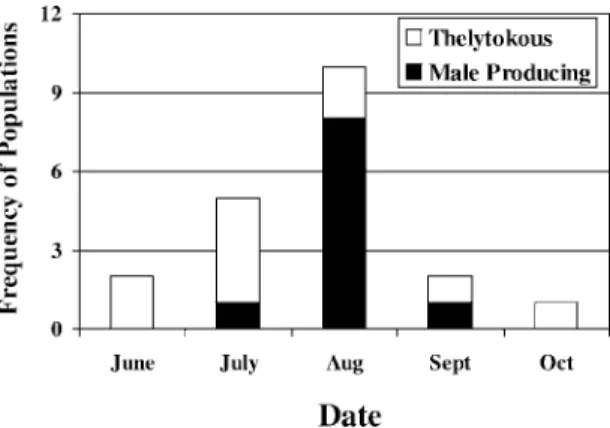
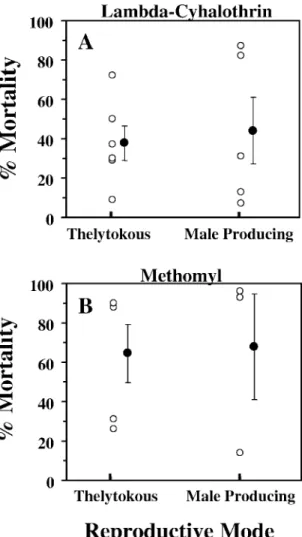
Figure
Documents relatifs
