Ultrastructural changes in chemically induced preneoplastic focal lesions in the rat liver: a stereological study
Texte intégral
Figure
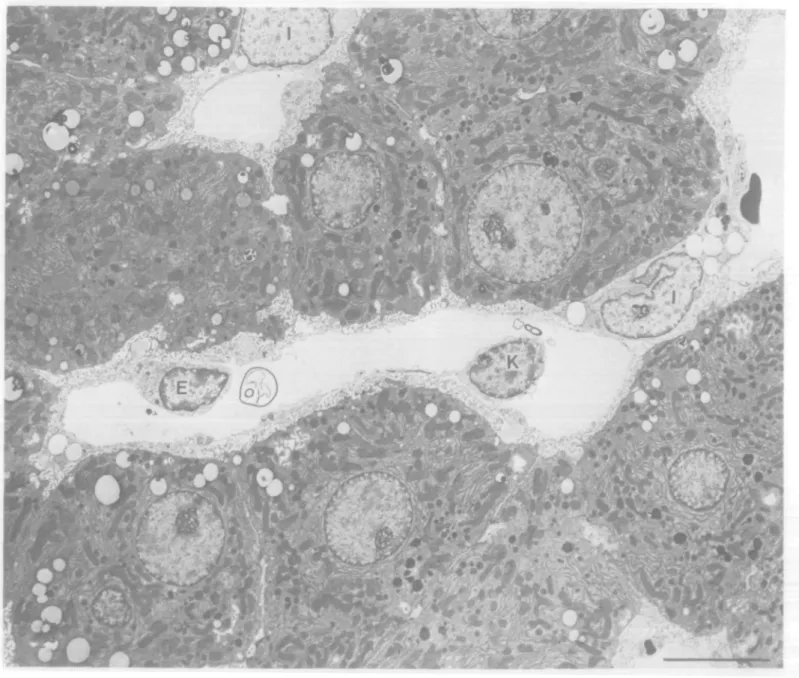
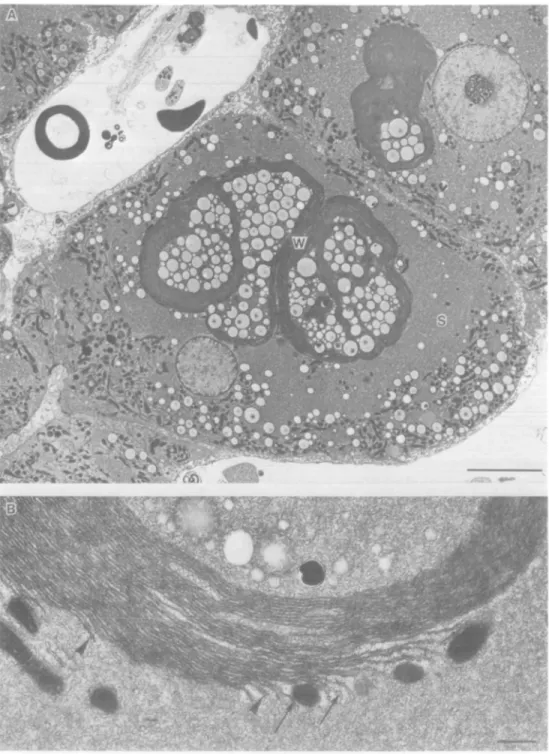

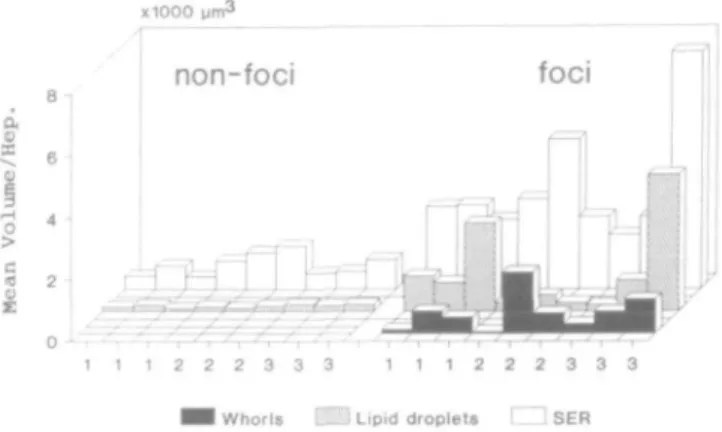
Documents relatifs
The XSL algorithm also defines a G-DEVS atomic model for each resource, task, stock and controller by a XML G-DEVS description. In order to illustrate the XML
The resonance states for the associated Floquet operators are described in Section 5 and in Section 6 we show that the Fermi golden rule holds for a generic class of time
Detecting the effect of land use changes on the flood regimes in the Mella River catchment requires information on the permeability of soils and rocks to implement a realistic model
return levels based on model projected mean and variance at any desired future
For this study the framework considered to characterise number sense is composed by the following compo- nents: (1) understand the meaning of numbers; (2) rec- ognise the relative
The fact that from λ A = 1.64 the volume variation does not decrease as in natural rubber indicates that, even though the elongation at crystallization is lower than in natural
Section 3 is devoted to the proof of the main result whereas in Section 4, we state and prove a sort of Poincaré-Wirtinger inequality related to the functional inequality proved
The goal of the present study was to provide quantita- tive information about the postnatal cellular maturation of the rat amygdala. Our major findings are as follows: 1)
