THE GABAB RECEPTOR AGONISTS BACLOFEN AND CGP 44532 PREVENT ACQUISITION OF ALCOHOL DRINKING BEHAVIOUR IN ALCOHOL-PREFERRING RATS
5
0
0
Texte intégral
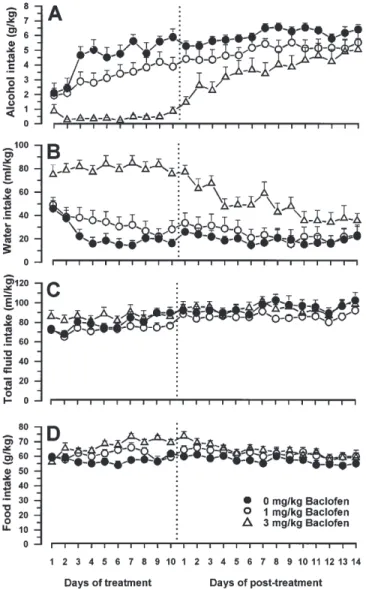
(2) 500. G. COLOMBO et al.. of human alcoholism. To further characterize the anti-alcohol effect of stimulation of the GABAB receptor, the present investigation was designed to assess the effect of baclofen and CGP 44532 on the acquisition of alcohol drinking behaviour in sP rats, that is a model of development of the predisposition toward excessive alcohol consumption in individuals at genetic risk of alcoholism. To this end, the sP rats used in the present study were alcohol-naive at the time of the start of baclofen administration. MATERIALS AND METHODS. RESULTS The present study evaluated, in two separate experiments, the effect of the repeated administration of baclofen and CGP 44532 on the acquisition of alcohol drinking behaviour in sP rats. In both experiments, mean daily alcohol intake in salinetreated rats rose to 5–6 g/kg/day [i.e. the amount of alcohol usually consumed daily by sP rats (Agabio et al., 2000)] within 4–7 days. Treatment with baclofen significantly [Ftreatment(2,216) = 22.597, P < 0.000005] and dose-dependently reduced the acquisition of alcohol drinking (Fig. 1A); in the rat group treated. Animals Male sP rats, from the 51st generation and 75 days old at the start of the study, were used. Rats were derived from a population of sP rats which underwent Caesarean delivery at Charles River (Lyon, France) for production of Specific Pathogen Free individuals. Rats were individually housed in standard plastic cages [size: 425 × 266 × 150 (height) mm] with wood chip bedding. The animal facility was under an inverted 12-h light: 12-h dark cycle (lights on at 23:00), at a constant temperature of 22 ± 2°C and relative humidity of ~60%. Rats were extensively habituated to handling and intraperitoneal (i.p.) injection. Rats were alcohol-naive before the start of the study. Experimental procedure Throughout the duration of the experiment, alcohol was offered in a free-choice regimen between the alcohol solution (10% v/v in tap water) and tap water with unlimited access for 24 h/day. Bottles were refilled every day with fresh solution and their left–right positions interchanged at random to avoid development of position preference. Food pellets (MIL Morini, San Polo d’Enza, RE, Italy) were always available. Rats were divided into three groups (n = 9 each) in the baclofen experiment and into four groups (n = 7 each) in the CGP 44532 experiment. In each experiment, rat groups were matched for body weight. In both experiments, alcohol presentation was initiated at the start of the dark phase of day 1. Rats received their first treatment, baclofen (0, 1 and 3 mg/kg; Sigma Chemical Co., St Louis, MO, USA) or CGP 44532 (0, 0.1, 0.3 and 1 mg/kg; Novartis, Basel, Switzerland), 20–30 min before alcohol presentation. Each drug was dissolved in saline (injection volume: 2 ml/kg) and injected i.p. Drug administration was repeated once a day (20–30 min before lights off) for 10 consecutive days. Alcohol, water and food intakes were monitored by weighing the bottles and food pellets (0.1 g accuracy) once daily immediately before the start of the dark phase. Recording of daily alcohol, water and food intake was performed throughout the 10 days of treatment as well as for an additional 14 (baclofen experiment) and 11 (CGP 44532 experiment) days after termination of treatment. Statistical analysis Data concerning daily alcohol, water, total fluid (i.e. the sum of alcohol solution and water consumed) and food intake during the 10-day treatment period and the post-treatment period of each experiment were expressed in g/kg, ml/kg, ml/kg and g/kg, respectively, and were evaluated separately by two-way (drug treatment; day) analysis of variance with repeated measures on the factor day.. Fig. 1. Effect of the repeated administration of the γ-aminobutyric acid (GABA)B receptor agonist baclofen on the acquisition of alcohol drinking behaviour in Sardinian alcohol-preferring (sP) rats. Baclofen (0, 1 and 3 mg/kg, i.p.) was injected once daily (20–30 min before lights off) for 10 consecutive days. Alcohol (10%, v/v) and water were offered under the two-bottle, free-choice regimen with unlimited access for 24 h/day starting from the first day of treatment with baclofen. Food pellets were always available. Alcohol, water and food intakes were monitored once a day immediately before lights off. The dashed line indicates the completion of the 10-day treatment period and the start of the post-treatment period. Each point is the mean ± SEM for n = 9 subjects..
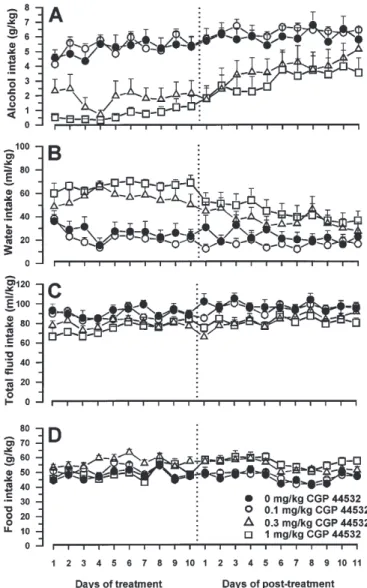
(3) GABAB RECEPTORS AND ALCOHOL DRINKING BEHAVIOUR. with 3 mg/kg baclofen, daily alcohol intake was virtually suppressed throughout the 10-day treatment period. Reduction in alcohol intake was associated with a compensatory increase in daily water intake [Ftreatment(2,216) = 27.544, P < 0.000001] (Fig. 1B), so that daily total fluid intake remained unchanged [Ftreatment(2,216) = 2.357, P > 0.05] (Fig. 1C). Daily food intake was significantly higher in the rat group treated with 3 mg/kg baclofen than in the saline-dosed control group [Ftreatment(2,216) = 7.854, P < 0.005] (Fig. 1D), probably because in the control group part of the total caloric intake was provided by alcohol. After treatment completion, daily alcohol intake in the 3 mg/kg baclofen-treated group increased progressively, reaching control values after ~2 weeks [Ftreatment(2,312) = 4.481, P < 0.05] (Fig. 1A). Consistently, water intake progressively diminished [Ftreatment(2,312) = 5.206, P < 0.05] (Fig. 1B). Daily total fluid intake [Ftreatment(2,312) = 1.188, P > 0.05] (Fig. 1C) and food intake [Ftreatment(2,312) = 2.651, P > 0.05] (Fig. 1D) did not differ among rat groups during the post-treatment period. Similarly, treatment with CGP 44532 resulted in a marked suppression of the acquisition of alcohol drinking behaviour [Ftreatment(3,216) = 13.992, P < 0.00005] (Fig. 2A); in the rat groups treated with 0.3 and 1 mg/kg CGP 44532, mean alcohol intake was steadily <2.5 and <1.5 g/kg/day, respectively, throughout the 10-day treatment period. Daily water intake was significantly [Ftreatment(3,216) = 13.156, P < 0.00005] increased in the rat groups treated with 0.3 and 1 mg/kg CGP 44532 (Fig. 2B). Total fluid intake [Ftreatment(3,216) = 4.113, P < 0.05] (Fig. 2C) as well as food intake [Ftreatment(3,216) = 3.301, P < 0.05] (Fig. 2D) were minimally affected by treatment with CGP 44532. After treatment completion, 11 days elapsed before alcohol intake in the 0.3 and 1 mg/kg CGP 44532-dosed groups approached the intake recorded in vehicle-treated control rats [Ftreatment(3,240) = 3.049, P < 0.05] (Fig. 2A). During the post-treatment period, water intake progressively decreased [Ftreatment(3,240) = 2.712, P > 0.05] (Fig. 2B), while total fluid intake did not differ among the rat groups [Ftreatment(3,240) = 2.076, P > 0.05] (Fig. 2C). Food intake was higher in the 0.3 and 1 mg/kg CGP 44532-treated groups [Ftreatment(3,240) = 5.961, P < 0.01] (Fig. 2D), presumably as a compensation of the lower caloric intake deriving from alcohol. DISCUSSION The results of the present study demonstrated that repeated daily administration of the GABAB receptor agonists, baclofen and CGP 44532, prevented the acquisition of alcohol drinking behaviour in selectively bred alcohol-preferring sP rats. Specifically, the daily average alcohol intake in the rat groups treated with 3 mg/kg baclofen and 1 mg/kg CGP 44532 was steadily <1 and <1.5 g/kg, respectively, throughout the 10-day treatment period. These results suggest that stimulation of the GABAB receptor results in a virtually complete blockade of the disclosure and experience of alcohol reinforcing properties, which is otherwise a rather rapid phenomenon in sP rats, as indicated by intakes of pharmacologically relevant amounts of alcohol from the very beginning of alcohol exposure in salinetreated rat groups. The doses of 3 mg/kg baclofen (corresponding to 14 µmol/kg) and 1 mg/kg CGP 44532 (corresponding. 501. Fig. 2. Effect of the repeated administration of the γ-aminobutyric acid (GABA)B receptor agonist CGP 44532 on the acquisition of alcohol drinking behaviour in Sardinian alcohol-preferring (sP) rats. CGP 44532 (0, 0.1, 0.3 and 1 mg/kg, i.p.) was injected once daily (20–30 min before lights off) for 10 consecutive days. Alcohol (10%, v/v) and water were offered under the two-bottle, free-choice regimen with unlimited access for 24 h/day starting from the first day of treatment with CGP 44532. Food pellets were always available. Alcohol, water and food intakes were monitored once a day immediately before lights off. The dashed line indicates the completion of the 10-day treatment period and the start of the post-treatment period. Each point is the mean ± SEM for n = 7 subjects.. to 6.5 µmol/kg) displayed comparable efficacy; the relatively higher potency of CGP 44532, compared to that of baclofen, in blocking the acquisition of alcohol drinking in sP rats is consistent with its higher binding affinity for the GABAB receptor (Froestl et al., 1995). The results of the present study extend to the acquisition phase the ability of baclofen to reduce alcohol intake in the so-called ‘maintenance’ model, i.e. in rats that were alcoholexperienced before the start of the baclofen treatment (Colombo et al., 2000). It should be noted, however, that the effect of baclofen on voluntary alcohol intake in rats is a.
(4) 502. G. COLOMBO et al.. matter of debate, as studies conducted so far have yielded equivocal results; indeed, baclofen has been found to reduce (Daoust et al., 1987; Colombo et al., 2000), produce no change on (Tomkins and Fletcher, 1996), or even to stimulate (Smith et al., 1992, 1999) alcohol intake in rats given a choice between an alcohol solution and water. Differences in the rat strain (unselected or selectively bred alcohol-preferring rats), baclofen dose-range, route of baclofen administration, and procedure of alcohol presentation may explain, at least in part, these discrepancies. With regard to the acquisition phase, Smith et al. (1992) reported that baclofen, administered at the single dose of 10 mg/kg, stimulated — although not specifically — the acquisition of alcohol intake in Long–Evans rats offered a choice between water and an alcohol solution in an ascending series of concentrations (2–10%) every other day. Again, several differences in the experimental procedures, including particularly the use of unselected Long Evans rats (presumably not possessing any particular innate predisposition to consume alcohol) or of genetically selected alcohol-preferring sP rats, might be responsible for the opposite outcomes of the study by Smith et al. (1992) and the present study. The cellular mechanism by which GABAB receptor agonists exert their reducing effect on alcohol intake has yet to be defined. Different lines of experimental evidence suggest a role for the mesolimbic dopamine system in the regulation of the positive reinforcing properties of alcohol. Accordingly, administration of low to moderate doses of alcohol have been reported to activate the firing rate of mesolimbic dopamine neurons (Gessa et al., 1985) and stimulate dopamine release in the nucleus accumbens (e.g. Imperato and Di Chiara, 1986; Weiss et al., 1993) in rats. Further, pharmacological manipulation of dopamine D1 and D2 receptor subtypes has been found to influence alcohol drinking behaviour in rodents and humans (e.g. Pfeffer and Samson, 1988; Enggasser and de Wit, 2001). GABAB receptors located in the ventral tegmental area have been suggested to contribute to the control of mesolimbic dopamine neurons, exerting — when stimulated — an inhibitory action (Kalivas, 1993; Yoshida et al., 1994; Westerink et al., 1996). A recent, preliminary microdialysis study from this laboratory demonstrated that acute administration of doses of baclofen, within the dose-range that reduces alcohol intake in rats, inhibits the alcohol-induced stimulation of dopamine release in the rat nucleus accumbens (Carta et al., 2001). Accordingly, baclofen has repeatedly been reported to suppress the stimulation of motor activity [a phenomenon mediated by activation of the mesolimbic dopamine system (see Wise and Bozarth, 1987)] induced by alcohol in mice (Cott et al., 1976; Humeniuk et al., 1993; Shen et al., 1998; Broadbent and Harless, 1999; Chester and Cunningham, 1999). Thus, the reducing effect of GABAB receptor agonists on voluntary alcohol intake might be the consequence of their ability to diminish the reinforcing properties of alcohol by inhibiting alcohol-stimulated dopamine release in the mesolimbic system. With regard to the results of the present study, it might be hypothesized that baclofen and CGP 44532 blocked the acquisition of alcohol drinking behaviour preventing the disclosure of the dopamine-mediated, positive reinforcing properties of alcohol, rendering the alcohol solution modestly attractive to the rat. In conclusion, the results of the present study demonstrate that the GABAB receptor agonists baclofen and CGP 44532. prevent with comparable efficacy the acquisition of alcohol drinking behaviour in alcohol-preferring sP rats, and suggest the involvement of the GABAB receptor in the mechanisms underlying the disclosure and experience of the psychopharmacological effects of alcohol that sustain voluntary alcohol drinking in this rat line. Acknowledgements — The authors are grateful to Mrs M. Elena Vincis, Mrs Gabriella Manca and Mr Antonio Pilleri, from Neuroscienze S.c.a r.l., for animal breeding and care, and Mrs Anne Farmer for language editing of the manuscript. The present study was partially supported by the European Community, the Italian Government and the Regione Autonoma della Sardegna through ‘Programma Operativo Plurifondo — Sardegna’.. REFERENCES Addolorato, G., Caputo, F., Capristo, E., Domenicali, M., Bernardi, M., Janiri, L., Agabio, R., Colombo, G., Gessa, G. L. and Gasbarrini, G. (2002a) Baclofen efficacy in reducing alcohol craving and intake — a double blind randomized controlled study. Alcohol and Alcoholism 37, 504–508. Addolorato, G., Caputo, F., Capristo, E., Janiri, L., Bernardi, M., Agabio, R., Colombo, G., Gessa, G. L. and Gasbarrini, G. (2002b) Rapid suppression of alcohol withdrawal syndrome by baclofen. American Journal of Medicine 112, 226–229. Agabio, R., Carai, M. A. M., Lobina, C., Pani, M., Reali, R., Bourov, I., Gessa, G. L. and Colombo, G. (2000) Dissociation of ethanol and saccharin preference in sP and sNP rats. Alcoholism: Clinical and Experimental Research 24, 24–29. Ahmadi-Abhari, S. A., Akhondzadeh, S., Assadi, S. M., Shabestari, O. L., Farzanehgan, Z. M. and Kamlipour, A. (2001) Baclofen versus clonidine in the treatment of opiates withdrawal, side-effects aspect: a double-blind randomized controlled trial. Journal of Clinical Pharmacy and Therapeutics 26, 67–71. Akhondzadeh, S., Ahmadi-Abhari, S. A., Assadi, S. M., Shabestari, O. L., Kashani, A. R. and Farzanehgan, Z. M. (2000) Double-blind randomized controlled trial of baclofen vs. clonidine in the treatment of opiates withdrawal. Journal of Clinical Pharmacy and Therapeutics 25, 347–353. Andrews, N. and File, S. E. (1993) Increased 5-HT release mediates the anxiogenic response during benzodiazepine withdrawal: a review of supporting neurochemical and behavioural evidence. Psychopharmacology 112, 21–25. Benedito, M. A. C. and Liete, J. R. (1981) Baclofen as an anticonvulsant in experimental models of convulsions. Experimental Neurology 72, 346–351. Bexis, S., Ong, J. and White, J. (2001) Attenuation of morphine withdrawal signs by the GABAB receptor agonist baclofen. Life Sciences 70, 395–401. Brebner, K., Froestl, W., Andrews, M., Phelan, R. and Roberts, D. C. S. (1999) The GABAB agonist CGP 44532 decreases cocaine selfadministration in rats: demonstration using a progressive ratio and a discrete trials procedure. Neuropharmacology 38, 1797–1804. Brebner, K., Childress, A. R. and Roberts, D. C. S. (2002a) A potential role for GABAB agonists in the treatment of psychostimulant addiction. Alcohol and Alcoholism 37, 478–484. Brebner, K., Froestl, W. and Roberts, D. C. S. (2002b) The GABAB antagonist CGP56433A attenuates the effect of baclofen on cocaine but not heroin self-administration in the rat. Psychopharmacology 160, 49–55. Broadbent, J. and Harless, W. E. (1999) Differential effects of GABAA and GABAB agonists on sensitization to the locomotor stimulant effects of ethanol in DBA/2 J mice. Psychopharmacology 141, 197–205. Carta, G., Satta, R., Pani, L., Colombo, G., Gessa, G. L. and Nava, F. (2001) Baclofen suppression of alcohol-induced dopamine release in the nucleus accumbens. Pharmacological Research 43 (Suppl. A), 35. Chester, J. A. and Cunningham, C. L. (1999) Baclofen alters ethanolstimulated activity but not conditioned place preference or taste aversion in mice. Pharmacology, Biochemistry and Behavior 63, 325–331..
(5) GABAB RECEPTORS AND ALCOHOL DRINKING BEHAVIOUR Colombo, G., Agabio, R., Carai, M. A. M., Lobina, C., Pani, M., Reali, R., Addolorato, G. and Gessa, G. L. (2000) Ability of baclofen in reducing alcohol intake and withdrawal severity: I — Preclinical evidence. Alcoholism: Clinical and Experimental Research 24, 58–66. Corrigall, W. A., Coen, K. M., Adamson, K. L., Chow, B. L. C. and Zhang, J. (2000) Response of nicotine self-administration in the rat to manipulations of mu-opioid and γ-aminobutyric acid receptors in the ventral tegmental area. Psychopharmacology 149, 107–114. Corrigall, W. A., Coen, K. M., Zhang, J. and Adamson, K. L. (2001) GABA mechanisms in the pedunculopontine tegmental nucleus influence particular aspects of nicotine self-administration selectively in the rat. Psychopharmacology 158, 190–197. Cott, J., Carlsson, A., Engel, J. and Lindqvist, M. (1976) Suppression of ethanol-induced locomotor stimulation by GABA-like drugs. Naunyn-Schmiedeberg’s Archives of Pharmacology 295, 203–209. Daoust, M., Saligaut, C., Lhuintre, J. P., Moore, N., Flipo, J. L. and Boismare, F. (1987) GABA transmission, but not benzodiazepine receptor stimulation, modulates ethanol intake by rats. Alcohol 4, 469–472. Diaz, S. L., Kemmling, A. K., Rubio, M. C. and Balerio, G. N. (2001) Lack of sex-related differences in the prevention by baclofen of the morphine withdrawal syndrome in mice. Behavioural Pharmacology 12, 75–79. Enggasser, J. L. and de Wit, H. (2001) Haloperidol reduces stimulant and reinforcing effects of ethanol in social drinkers. Alcoholism: Clinical and Experimental Research 25, 1448–1456. Fattore, L., Cossu, G., Martellotta, M. C., Deiana, S. and Fratta, W. (2001) Baclofen antagonises intravenous self-administration of gamma-hydroxybutyric acid in mice. Neuroreport 12, 2243–2246. Fattore, L., Cossu, G., Martellotta, M. C. and Fratta, W. (2002) Baclofen antagonises intravenous self-administration of nicotine in mice and rats. Alcohol and Alcoholism 37, 495–498. File, S. E., Zharkovsky, A. and Gulati, K. (1991) Effects of baclofen and nitrendipine on ethanol withdrawal responses in the rat. Neuropharmacology 30, 183–190. Froestl, W., Mickel, S. J., Hall, R. G., von Sprecher, G., Strub, D., Baumann, P. A., Brugger, F., Gentsch, C., Jaekel, J., Olpe, H.-R., Rihs, G., Vassout, A., Waldmeier, P. C. and Bittiger, H. (1995) Phosphinic acid analogues of GABA. 1. New potent and selective GABAB agonists. Journal of Medicinal Chemistry 38, 3297–3312. Gessa, G. L., Muntoni, F., Collu, M., Vargiu, L. and Mereu, G. (1985) Low doses of ethanol activate dopaminergic neurons in the ventral tegmental area. Brain Research 348, 201–203. Humeniuk, R. E., White, J. M. and Ong, J. (1993) The role of GABAB receptors in mediating the stimulatory effects of ethanol in mice. Psychopharmacology 111, 219–224. Humeniuk, R. E., White, J. M. and Ong, J. (1994) The effects of GABAB ligands on alcohol withdrawal in mice. Pharmacology, Biochemistry and Behavior 49, 561–566. Imperato, A. and Di Chiara, G. (1986) Preferential stimulation of dopamine release in the nucleus accumbens of freely moving rats by ethanol. Journal of Pharmacology and Experimental Therapeutics 239, 219–228. Kalivas, P. W. (1993) Neurotransmitter regulation of dopamine neurons in the ventral tegmental area. Brain Research Reviews 18, 75–113. Kalivas, P. W., Duffy, P. and Eberhardt, H. (1990) Modulation of A10 dopamine neurons by γ-aminobutyric acid agonists. Journal of Pharmacology and Experimental Therapeutics 253, 858–866. Kemmling, A. K., Rubio, M. C. and Balerio, G. N. (2002) Baclofen prevents morphine withdrawal irrespective of seasonal variation. Behavioural Pharmacology 13, 87–92. Leite-Morris, K. A., Fukudome, E. Y. and Kaplan, G. B. (2002) Opiateinduced motor stimulation is regulated by γ-aminobutyric acid type B receptors found in the ventral tegmental area in mice. Neuroscience Letters 317, 119–122. Li, S.-M., Yin, L.-L., Ren, Y.-H., Pan, L.-S. and Zheng, J.-W. (2001) GABAB receptor agonist baclofen attenuates the development and expression of d-methamphetamine-induced place preference in rats. Life Sciences 70, 349–356.. 503. Ling, W., Shoptaw, S. and Majewska, D. (1998) Baclofen as a cocaine anti-craving medication: a preliminary clinical study. Neuropsychopharmacology 18, 403–404. Menon, M. K., Clark, W. G. and Vivonia, C. (1980) Interaction between phencyclidine (PCP) and gaba-ergic drugs: clinical implications. Pharmacology, Biochemistry and Behavior 12, 113–117. Paterson, N. E., Markou, A. and Froestl, W. (2001) Nicotine selfadministration in rats on a progressive ratio schedule: administration of the GABAB agonist CGP-44532 decreased breaking points. Society for Neurosciences Abstract Book 27, 1774. Petry, N. M. (1997) Benzodiazepine-GABA modulation of concurrent ethanol and sucrose reinforcement in the rat. Experimental and Clinical Psychopharmacology 5, 183–194. Pfeffer, A. O. and Samson, H. H. (1988) Haloperidol and apomorphine effects on ethanol reinforcement in free feeding rats. Pharmacology, Biochemistry and Behavior 29, 343–350. Phillis, B. D., Ong, J., White, J. M. and Bonnielle, C. (2001) Modification of d-amphetamine-induced responses by baclofen in rats. Psychopharmacology 153, 277–284. Sandoval, M. R. L. and Palermo-Neto, J. (1985) Central nervous system supersensitivity and withdrawal from long-term treatment with barbital. Neuropharmacology 24, 1247–1251. Shen, E. H., Dorow, J., Harland, R., Burkhart-Kasch, S. and Phillips, T. J. (1998) Seizure sensitivity and GABAergic modulation of ethanol sensitivity in selectively bred FAST and SLOW mouse lines. Journal of Pharmacology and Experimental Therapeutics 287, 606–615. Smith, B. R., Robidoux, J. and Amit, Z. (1992) GABAergic involvement in the acquisition of voluntary ethanol intake in laboratory rats. Alcohol and Alcoholism 27, 227–231. Smith, B. R., Boyle, A. E. L. and Amit, Z. (1999) The effects of GABAB agonist baclofen on the temporal and structural characteristics of ethanol intake. Alcohol 17, 231–240. Tarika, J. S. and Winger, G. (1980) The effects of ethanol, phenobarbital, and baclofen on ethanol withdrawal in the rhesus monkey. Psychopharmacology 70, 201–208. Tomkins, D. M. and Fletcher, P. J. (1996) Evidence that GABAA but not GABAB receptor activation in the dorsal raphe nucleus modulates ethanol intake in Wistar rats. Behavioural Pharmacology 7, 85–93. Tsuji, M., Nakagawa, Y., Ishibashi, Y., Yoshii, T., Takashima, T., Shimada, M. and Suzuki, T. (1996) Activation of ventral tegmental GABAB receptors inhibits morphine-induced place preference in rats. European Journal of Pharmacology 313, 169–173. Weiss, F., Lorang, M. T., Bloom, F. E. and Koob, G. F. (1993) Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. Journal of Pharmacology and Experimental Therapeutics 267, 250–258. Westerink, B. H. C., Kwint, H.-F. and deVries, J. B. (1996) The pharmacology of mesolimbic dopamine neurons: a dual-probe microdialysis study in the ventral tegmental area and nucleus accumbens of the rat brain. Journal of Neuroscience 16, 2605–2611. Wise, R. A. and Bozarth, M. A. (1987) A psychomotor stimulant theory of addiction. Psychological Reviews 94, 469–492. Woo, S.-H., Kim, H.-S., Yun, J.-S., Lee, M.-K., Oh, K.-W., Seong, Y.-H., Oh, S.-K. and Jang, C.-G. (2001) Inhibition of baclofen on morphine-induced hyperactivity, reverse tolerance and postsynaptic dopamine receptor supersensitivity. Pharmacological Research 43, 335–340. Xi, Z.-X. and Stein, E. A. (2002) GABAergic mechanisms of opiate reinforcement. Alcohol and Alcoholism 37, 485–494. Yoshida, M., Yokoo, H., Tanaka, T., Emoto, H. and Tanaka, M. (1994) Opposite changes in the mesolimbic dopamine metabolism in the nerve terminal and cell body sites induced by locally infused baclofen in the rat. Brain Research 636, 111–114. Zarrindast, M.-R. and Mausa-Ahmadi, E. (1999) Effects of GABAergic system on naloxone-induced jumping in morphine-dependent mice. European Journal of Pharmacology 381, 129–133..
(6)
Figure
Documents relatifs
Plasma alcohol concentrations were determined in H3R KO and WT mice after acute alcohol administration (2.0 g/kg, at 5 min) and following 4-hour drinking in the dark session.. In
For hand rub 1, the total absorbed doses, calculated from ethanol with an inhalation flow of 24 L/min and an absorption rate of 62%, were 46.5 mg after one hygienic hand
However, no study has assessed the effects of chronic intermittent exposure to ethanol vapor on bone mineral density and bone microarchitecture.. This was the objective of
Le pavage fonctionne-t-il encore si le quadrilatère ABCD possède un angle rentrant comme la figure ci-contre.. (On dit dans ce cas que le quadrilatère
In particular, a theoretical explanation is given for the observation that in Europe many firms borrow from several banks while one bank takes the role of a relationship bank to
As there is no specific questionnaire to assess sugar appetence and craving, we used the following tools on D1, D15 and D45: (1) a visual analogue scale with scores ranging from 0
School -based education and mass media information campaigns are the most widely used approaches in the primary prevention of alcohol - related problems.. Available research
A more realistic figure is that provided by Powell (unpublished data, 1990) who reviewed a number of studies and concluded that fewer than 20% of firms in the United Kingdom had
