Comparative localization of serotonin-like immunoreactive cells in Thaliacea informs tunicate phylogeny
13
0
0
Texte intégral
Figure
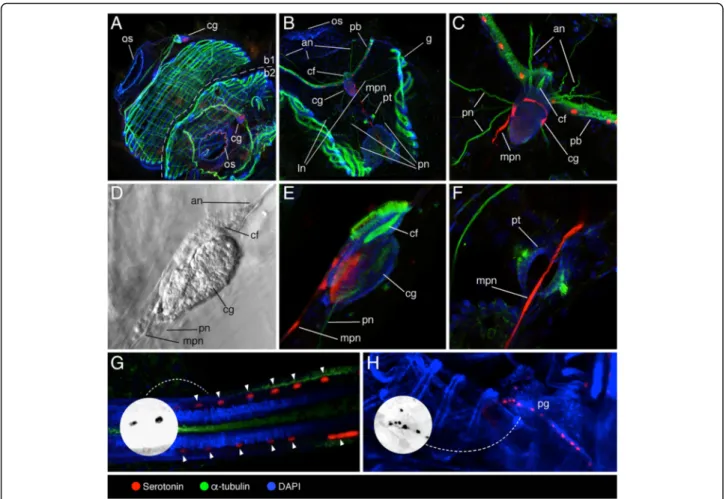
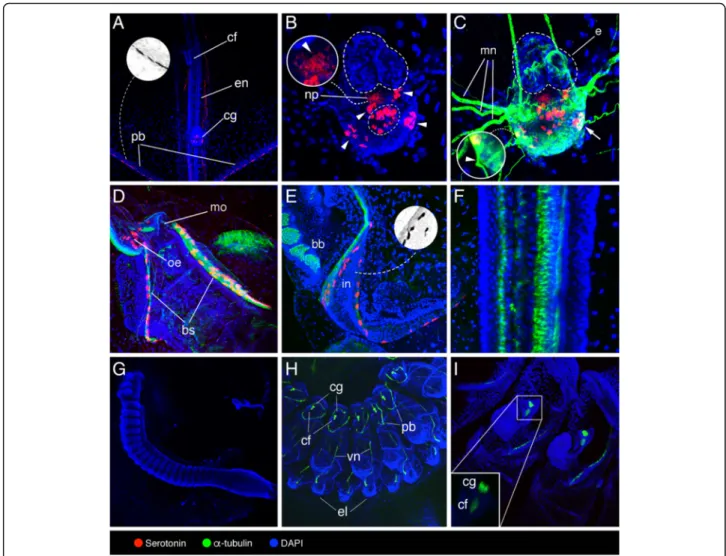
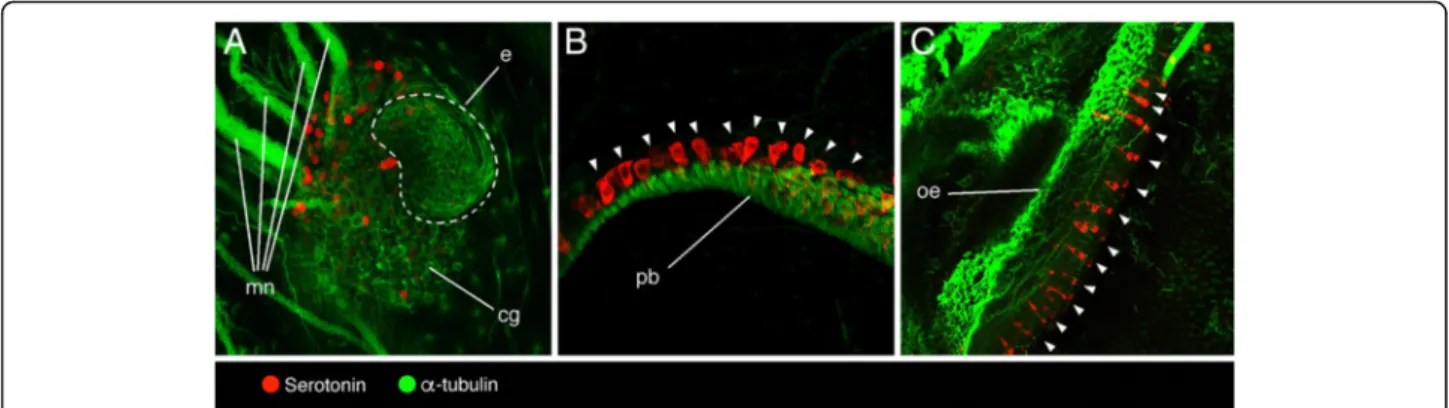
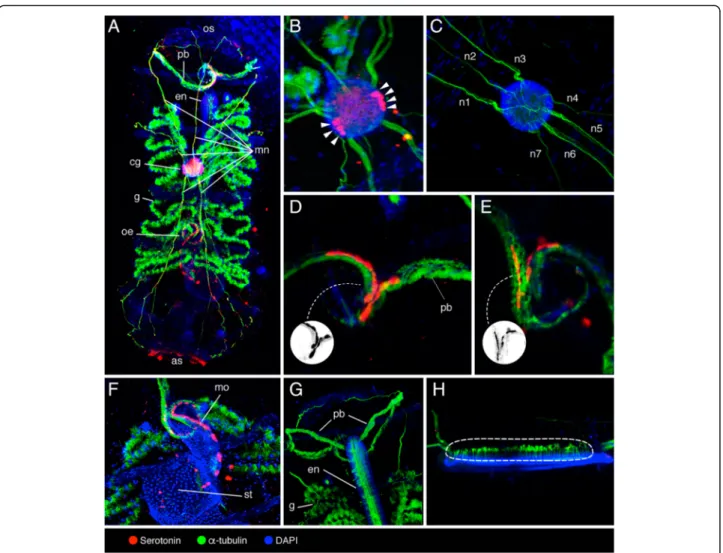
+4
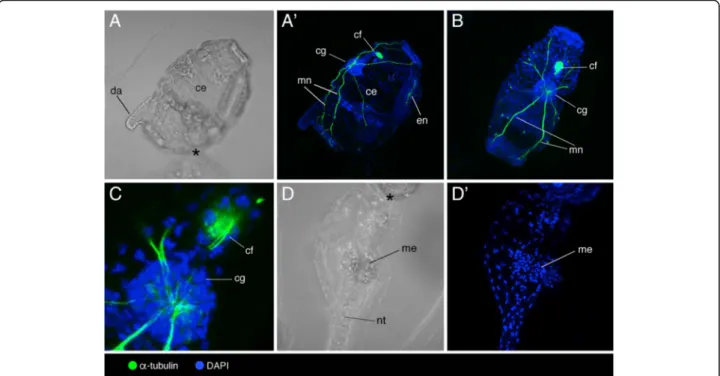
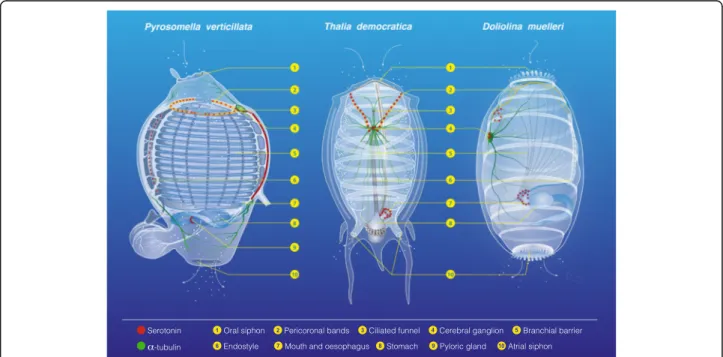
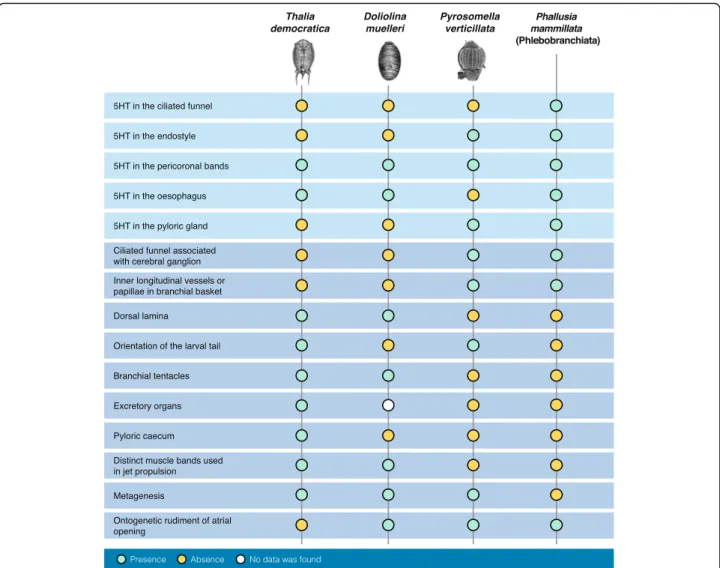
Documents relatifs