HAL Id: hal-01308042
https://hal.archives-ouvertes.fr/hal-01308042
Submitted on 27 Apr 2016
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Large-scale simulation of steady and time-dependent
active suspensions with the force-coupling method
Blaise Delmotte, Eric Keaveny, Franck Plouraboué, Éric Climent
To cite this version:
Blaise Delmotte, Eric Keaveny, Franck Plouraboué, Éric Climent. Large-scale simulation of steady
and time-dependent active suspensions with the force-coupling method. Journal of Computational
Physics, Elsevier, 2015, 302, pp.524-547. �10.1016/j.jcp.2015.09.020�. �hal-01308042�
O
pen
A
rchive
T
OULOUSE
A
rchive
O
uverte (
OATAO
)
OATAO is an open access repository that collects the work of Toulouse researchers and
makes it freely available over the web where possible.
This is an author-deposited version published in :
http://oatao.univ-toulouse.fr/
Eprints ID : 14290
To link to this article: DOI :
10.1016/j.jcp.2015.09.020
URL :
http://dx.doi.org/10.1016/j.jcp.2015.09.020
To cite this version : Delmotte, Blaise and Keaveny, Eric and
Plouraboué, Franck and Climent, Eric
Large-scale simulation of steady
and time-dependent active suspensions with the force-coupling method
.
(2015) Journal of Computational Physics, vol. 302. pp. 524-547. ISSN
0021-9991
Any correspondance concerning this service should be sent to the repository
administrator:
[email protected]
Large-scale
simulation
of
steady
and
time-dependent
active
suspensions
with
the
force-coupling
method
Blaise Delmotte
a,
b,
∗
,
Eric
E. Keaveny
c,
∗∗
,
Franck Plouraboué
a,
b,
Eric Climent
a,
baUniversityofToulouse,INPT-UPS,InstitutdeMécaniquedesFluides,Toulouse,France bIMFT,CNRS,UMR5502, 1AlléeduProfesseurCamilleSoula,31400Toulouse,France
cDepartmentofMathematics,ImperialCollegeLondon,SouthKensingtonCampus,London,SW72AZ,UK
a
b
s
t
r
a
c
t
Keywords:
Forcecouplingmethod LowReynoldsnumber Activesuspension Swimminggait Collectivedynamics Highperformancecomputing
We present a new development of the force-coupling method (FCM) to address the accuratesimulation of alarge number of interactingmicro-swimmers. Ourapproach is based on the squirmer model, which we adapt to the FCM framework, resulting in a methodthat is suitable for simulating semi-dilute squirmer suspensions. Other effects, such as steric interactions, are considered with our model. We test our method by comparing the velocity field around a single squirmer and the pairwise interactions betweentwosquirmerswithexactsolutionstotheStokesequationsandresultsgivenby othernumerical methods.We alsoillustrate ourmethod’sability to describespheroidal swimmer shapes and biologically-relevant time-dependent swimming gaits. We detail the numerical algorithm used to compute the hydrodynamic coupling between a large collection (104–105) of micro-swimmers. Using this methodology, we investigate the
emergenceofpolarorderinasuspensionofsquirmersandshowthatforlargedomains, both the steady-state polar order parameter and the growth rate of instability are independentofsystemsize.Theseresultsdemonstratetheeffectivenessofourapproach toachievenearcontinuum-levelresults,allowingforbettercomparisonwithexperimental measurementswhilecomplementingandinformingcontinuummodels.
1. Introduction
Suspensionsofactive,self-propelledparticles arise inbothbiological systems,such aspopulationsofmicro-organisms
[1–4]andsynthetic,colloidalsystems[5].Thesesuspensionscanexhibit theformationofcoherentstructuresandcomplex flow patterns which may lead to enhanced mixing of chemicals in the surrounding fluid, the alteration of suspension rheology,or,inthebiologicalcase,increasednutrientuptakebyapopulationofmicro-organisms.Inadditiontopromising applicationssuchasalgaebiofuels[6,7],characterizingthecollectivedynamicsfoundinthesesuspensionsisoffundamental importancetounderstandingzooplanktondynamics[8,9]andmammalfertility[10,11].
Themathematicalmodelingofactivesuspensionsentailsdescribinghowindividualswimmersmoveandinteractin re-sponseto theflow fieldsthat theygenerate[12–14].Itis particularlyimportantforthesemodelsto be abletohandlea largecollectionofswimmersinorderto obtainsuspension propertiesatthe lab/insitu scale.The modelingofthe
collec-*
Correspondingauthorat:IMFT,2alléeduProfesseurCamilleSoula,31400Toulouse.**
Correspondingauthor.E-mailaddresses:[email protected](B. Delmotte),[email protected](E.E. Keaveny),[email protected](F. Plouraboué),
[email protected](E. Climent).
tive behaviorofactive matterhasbeenavibrantarea ofresearchduringthelast decade[15,16,14,17],tociteonly afew recent reviews. Generallyspeaking, themodeling approaches canbe sorted into two categories:continuum theories and particle-based simulations.Most ofthe continuum modelsare generallyvalid fordilute suspensionswherethe hydrody-namicdisturbancesaregivenbyamean-fielddescriptionoffar-fieldhydrodynamicinteractions[18,19,16].Recentadvances towards more concentrated suspensionsinclude steric interactions [20], butthe inclusion ofhigh-order singularities due to particlesizeremains outstanding.Despitethis, thesemodels arevery attractiveastheynaturally provideadescription ofthedynamicsatthepopulationlevelandtheresultingequationscanbeanalyzed using awiderangeofanalyticaland numericaltechniques.
Particle-basedsimulations resolvethedynamicsofeachindividualswimmer andfromtheir positionsandorientations, construct a picture of the dynamics of the suspension as a whole. As discussed in [16], particle-based models provide opportunitiesto (i)test continuum theories,(ii) analyze finite-sizeeffectsresultingfromadiscrete numberofswimmers, (iii)exploremorecomplexinteractionsbetweenswimmersand/or boundaries,andinsomecases,(iv)revealtheeffectsof short-rangehydrodynamicinteractionsand/orstericrepulsion.Variousmodelshavebeenproposedinthiscontext,each us-ingdifferentapproximationstoaddressthedifficultproblemsofresolvingthehydrodynamicinteractionsandincorporating the geometryof the swimmers.Some of thefirst such models used point force distributions to createdumbbell-shaped swimmers [21–23],slender-body theory to modela slipvelocity along thesurfaces ofrod-likeswimmers [24,25],orthe squirmer model[26,27]to examinethe interactions betweensphericalswimmers[28].Theseinitial studiesprovided im-portantfundamentalresultsconnectingthepropertiesoftheindividualswimmerstotheemergenceofcollectivedynamics. Basedontheirsuccess,thesemodelshavebeenmorerecentlyincorporatedintoanumberofnumericalapproachesfor sus-pensionandfluid-structureinteractionsimulationsincludingStokesiandynamics[29–31],theimmersedboundarymethod
[32,33],LatticeBoltzmannmethods[34,35],andhybridfiniteelement/penalizationschemes[36].Thishasallowedforboth increasedswimmer numbersaswell astheincorporationofother effectssuch asstericinteractions,external boundaries, andaligningtorques.
In thispaper,we introduce an extension ofthe force couplingmethod (FCM) [37,38],an approach forthe large-scale simulationofpassiveparticles,tocapturethemany-bodyinteractionsbetweenactiveparticles.FCMreliesonaregularized, ratherthanasingular,multipoleexpansiontoaccountforthehydrodynamicinteractionsbetweentheparticles.Itincludes a higher-ordercorrection duetoparticlerigidityby enforcingtheconstraintofzero-averagedstrain rateinthevicinity of eachparticle.Sincetheforcedistributionshavebeenregularized,thetotalparticleforce,includingthatassociatedwiththe constraints,canbeprojectedontoagridoverwhichthefluidflowcanbefoundnumerically.Thisallowsthehydrodynamic interactions forallparticles tobe resolvedsimultaneously. Thismeshcanbe structured andsimplesuch that anefficient parallelStokessolvercanbeusedtofindthehydrodynamicinteractions.
WeextendFCMtoactiveparticlesbyintroducingtheregularizedsingularitiesintheFCMmultipoleexpansionthathave adirectcorrespondencetothesurfacevelocitymodesofthesquirmermodel[27].Withthesetermsincluded,wethenrely ontheusual FCMframework toresolvethehydrodynamic interactionsinaveryefficientmanner.We showthatby using the full capacityafforded byFCM, we are ableto accurately simulate active particlesuspensionsin thesemi-dilute limit with O
(
104–105)
swimmers.Usingthismethod,we examinetheinfluenceofdomainsizeonthesteady-statepolarorderobservedforsquirmersuspensions.Atthesametime,weshowthatourmethodisquiteversatile,beingabletohandle time-dependentswimminggaits,ellipsoidalswimmershapes,andstericinteractions,eachataminimaladditionalcomputational cost.Weexploreindetailhowtoincorporatebiologically-relevant,time-dependentswimminggaitsbytuningourmodelto the recentmeasurements oftheoscillatory flow aroundChlamydomonasRheinardtii[39].These experimentsrevealedthat consideringtime-averagedflowsforsuchmicro-organismsmayoversimplifythehydrodynamicinteractionsbetween neigh-bors. Time-dependency isalsocloselyassociated withtheway zooplankton feed, mixthe surroundingfluid, andinteract witheachother [6,9].Asstatedin[40],modeling micro-swimmerswitha time-dependentswimminggaitmightbe more realistic andshould be includedinmathematical models andcomputersimulations. We show thattime-dependence can indeedaffecttheoverallorganizationofthesuspension.
Weorganizeourpaperasfollows:InSection2,wereviewFCMandpresentthetheoreticalbackgroundforitsadaptation to active particles. Section 3 details the numerical method, its algorithmic implementation and how the computational workscaleswiththeparticlenumber.InSection4,wevalidatethemethodandtestitsaccuracybycomparingflowfields, trajectories, andpairwise interactions withprevious resultsavailable inthe literature. The effectiveness of ourapproach isdemonstratedinSection 5wherewe presentresultsfromlarge-scalesimulations ofactiveparticlesuspensions.Finally, extensionsofFCMtomorecomplexscenariosareintroducedinSection6.Wesimulatesuspensionsofspheroidalswimmers and demonstratethe newimplementation oftime-dependentswimming gaits.Here, we also presentpreliminary results showingtheeffectoftime-dependenceonsuspensionproperties.
2. SquirmersusingFCM
The force-couplingmethod(FCM)developedbyMaxey andcollaborators[37,38]isan effectiveapproachforthe large-scalesimulationofparticulatesuspensions,especiallyformoderatelyconcentratedsuspensionsatlowReynoldsnumber.In thiscontext,ithasbeenusedtoaddressavarietyofproblemsinmicrofluidics[41],biofluiddynamics[42],andmicron-scale locomotion[43–45]. FCMhasalso beenextendedto incorporatefiniteReynoldsnumbereffects[46],thermalfluctuations
FCM has beenused to address questions infundamental fluid dynamicsin regimes whereinertial effects are important and/orthere isahighvolume fractionofparticles[49,51].At thesametime, FCM hasbeenusedtoaddress problemsof technologicalimportance,suchasmicro-bubbledragreduction[46]andthedynamicsofcolloidalparticles[47].Inthis sec-tion,weexpandon[52]anddevelopthetheoreticalunderpinningsofFCM’sfurtherextensiontoactiveparticlesuspensions usingthesquirmermodelproposedbyLighthill[26],advancedbyBlake[27],andemployedbyIshikawaetal.[28].Tobegin thispresentation,wegiveanoverviewofFCM,establishingalsothenotationthatwillbeusedthroughoutthepaper. 2.1. FCMforpassiveparticles
Considerasuspensionof Np rigidsphericalparticles,eachhavingradiusa.Eachparticlen,
(
n=
1,
. . . ,
Np)
,iscenteredatYn andsubject toforce Fn andtorque
τ
n.Todeterminetheir motionthroughthesurrounding fluid,wefirst representeachparticlebyaloworder,finite-forcemultipoleexpansionintheStokesequations
∇
p−
η
∇
2u=
X
n Fn1
n(
x)
+
1 2τ
n× ∇2
n(
x)
+
Sn· ∇2
n(
x)
∇
·
u=
0.
(1)InEq.(1),Sn aretheparticlestressletsdeterminedthroughaconstraintonthelocalrate-of-strainasdescribedbelow.Also
inEq.(1)arethetwoGaussianenvelopes,
1
n(
x)
= (
2π σ
12)
−3/2e−|x−Yn| 2/2σ2 1,
2
n(
x)
= (
2π σ
2 2)
−3/2e−|x−Yn| 2/2σ2 2,
(2)usedtoprojecttheparticleforcesontothefluid.
Aftersolving Eq.(1),thevelocity,Vn,angularvelocity,
Ä
n, andlocalrate-of-strain, En,ofeachparticlen arefound byvolumeaveragingoftheresultingfluidflow,
Vn
=
Z
u1
n(
x)
d3x (3)Ä
n=
1 2Z
[∇
×
u]2
n(
x)
d3x,
(4) En=
1 2Z h
∇
u+ (∇
u)
Ti
2
n(
x)
d3x,
(5)wheretheintegrationisperformedovertheentiredomain.InorderforEqs.(3)–(5)torecoverthecorrectmobilityrelations forasingle,isolated sphere,namelythat V
=
F/(
6π
aη
)
andÄ
=
τ
/(
8π
a3η
)
,theenvelopelengthscales needtobeσ
1
=
a
/
√
π
andσ
2=
a/
¡
6
√
π
¢1
/3.Astheparticlesarerigid,thestressletsarefoundbyenforcingtheconstraintthatEn=
0foreachparticlen[38]. 2.2.Squirmermodel
Inadditiontoundergoingrigidbodymotionintheabsenceofappliedforcesortorques,activeandself-propelled parti-clesarealsocharacterizedbytheflowstheygenerate.Tomodelsuchparticles,wewillneedtoincorporatetheseflowsinto FCM.We accomplish thisbyadapting theaxisymmetric squirmermodel[26–28] tothe FCM framework,though we note thatamoregeneralsquirmermodel[53] withnon-axysimmetricsurfacemotioncouldalsobeconsidered.
Thesquirmermodelconsistsofasphericallyshaped,self-propelledparticlethatutilizesaxisymmetricsurfacedistortions tomovethroughfluidwithspeedU inthedirectionp.Iftheamplitudeofthedistortionsissmallcomparedtotheradius, a,ofthesquirmer,theireffectcanberepresentedbythesurfacevelocity,v
(
r=
a)
=
vrrˆ
+
vθˆθ
wherevr
=
U cosθ
+
∞X
n=0 An(
t)
Pn(
cosθ ),
(6) vθ= −
U sinθ
−
∞X
n=1 Bn(
t)
Vn(
cosθ ).
(7)Here, Pn
(
x)
aretheLegendrepolynomials, Vn(
cosθ )
=
2
n
(
n+
1)
sinθ
P′
n
(
cosθ ),
(8)theangle
θ
ismeasured withrespect totheswimmingdirectionp,and Pn′(
x)
=
d Pn/
dx.Inorder forthesquirmerto beforce-free,wehave
U
=
1Fig. 1.Decomposition of squirmer velocity field forβ=1.
FollowingIshikawaetal.[28],we considerareducedsquirmermodelwhere An
=
0 foralln andBn=
0 foralln>
2.Wethereforeonly havethefirsttwo termsoftheseries.Forthiscase, theresultingflow fieldinthe framemoving withthe swimmerisgivenby ur
(
r, θ )
=
2 3B1 a3 r3P1(
cosθ )
+
µ
a4 r4−
a2 r2¶
B2P2(
cosθ )
(10) uθ(
r, θ )
=
1 3B1 a3 r3V1(
cosθ )
+
a4 r4B2V2(
cosθ )
(11)wherewehaveusedU
=
2B1/3 fromEq.(9).Intermsofp,x,andr,thisbecomesu
(
x)
= −
B1 3 a3 r3µ
I−
3xx T r2¶
p+
µ
a4 r4−
a2 r2¶
B2P2³
p·
x r´
x r−
3a 4 r4B2³
p·
x r´ µ
I−
xx T r2¶
p=
uB1+
uB2 (12)Fig. 1 shows an example of a flow field given by Eq. (12), as well as the flows uB1 and uB2 related to the B1 and B2
contributions. While we have already seen that B1 is related to the swimming speed, it can be shown [28] that B2 is
directlyrelatedtothestresslet
G
=
43
π η
a2
(
3pp−
I)
B2 (13)generated by the surface distortions. This termsets the leading-order flow field that decays like r−2.We can introduce theparameter
β
=
B2/
B1 whichdescribestherelativestressletstrength.Inaddition,ifβ >
0,thesquirmerbehaves likea ‘puller’,bringingfluidinalongpandexpellingitlaterally,whereasifβ <
0,thesquirmerisa‘pusher’,expellingfluidalong pandbringingitinlaterally.ToadaptthismodeltotheFCMframework,wefirstrecognizethattheflowgivenbyEq.(12)canberepresentedbythe followingsingularitysystemintheStokesequations
∇
p−
η
∇
2u=
G· ∇
µ
δ(
x)
+
a 2 6∇
2δ(
x)
¶
+
H∇
2δ(
x)
(14)∇
·
u=
0 (15)wherethedegeneratequadrupoleisrelatedtoB1 through
H
= −
43π η
a3B1p (16)andthestressletGisgivenbyEq.(13).Wecandrawaparallelbetweenthesesingularitiesandtheregularizedsingularities used withFCM. The stressletterminEq. (14)isthe gradient ofthe singularitysystemfor asingle spheresubject to an appliedforce.Accordingly,thecorrespondingregularizedsingularityinFCMis
∇1(
x)
,where1(
x)
isgivenbyEq.(2).Itis importanttonotethateventhoughwereplacetwosingularforce distributionswiththeoneregularizedFCM distribution, the particular choice of1(
x)
will yield flows that are asymptotic to both singular flow fields [37]. For the degenerate quadrupole,however,thereisnotacorrespondingnaturalchoicefortheregularizeddistribution.Following[54],wechooseaGaussianenvelopewithalength-scale smallenough toyieldan accuraterepresentationofthesingularflow, butnotso smallastosignificantlyincreasetheresolutionneededinanumericalsimulation(seeSection4.1).Wethereforeemploythe FCMenvelopefortheforcedipole andreplacethesingulardistributionby
∇
22(
x)
.Thus,forasinglesquirmer,theStokesequationswiththeFCMsquirmerforcedistributionare
∇
p−
η
∇
2u=
G· ∇1(
x)
+
H∇
22(
x)
∇
·
u=
0.
(17)AsweshowSection4,theflowsatisfyingEq.(17)closelymatchesthatobtainedusingtheoriginalsingularitydistribution, Eq.(14).
2.3.Squirmerinteractionsandmotion
UsingFCM,thetaskofcomputingtheinteractionsbetweensquirmersisrelativelystraightforward.WenowconsiderNp
independentsquirmerswhereeachsquirmerhasswimmingdipole Gn anddegenerate quadrupoleHn.Thesquirmersmay
alsobesubjecttoexternal forcesFn andtorques
τ
n.UsingthelinearityoftheStokesequations,wecombineEqs.(1)and(17)toobtain
∇
p−
η
∇
2u=
X
n Fn1
n(
x)
+
1 2τ
n× ∇2
n(
x)
+
Sn· ∇2
n(
x)
+
Gn· ∇1
n(
x)
+
Hn∇
22
n(
x)
(18)∇
·
u=
0 (19)fortheflowfieldgeneratedbythesuspension.
Afterfindingtheflowfield,wedeterminethemotionofthesquirmersusingEqs.(3)–(5)withtwomodifications.First, weneed toadd theswimmingvelocity,U pn,toEq.(3).Second,we mustsubtractthe artificial,self-induced velocityand
thelocalrate-of-strainduetothesquirmingmodes.Theself-inducedvelocityisgivenby
Wn
=
Z
A
·
Hn1(
x)
d3x (20)wherethetensorA,inindexnotation,is[37]
Ai j
(
x)
=
1 4π η
r3·
δ
i j−
3xixj r2¸
erfµ
rσ
2√
2¶
(21)−
1η
·³
δ
i j−
xixj r2´
+
µ
δ
i j−
3xixj r2¶ ³
σ
2 r´
2¸
2(
x).
(22)Theself-inducedrate-of-strainisgivenby
Kn
=
Z
1 2³
∇
R·
Gn+ (∇
R·
Gn)
T´
2(
x)
d3x (23)wheretheexpressionforthethirdranktensorRcanbe foundin[38].Takingtheseself-induced effectsintoaccount,the motionofasquirmern isgivenby
Vn
=
U pn−
Wn+
Z
u1
n(
x)
d3x (24)Ä
n=
1 2Z
[∇
×
u]2
n(
x)
d3x (25) En= −
Kn+
1 2Z h
∇
u+ (∇
u)
Ti
2
n(
x)
d3x.
(26)Asbefore,thestressletsSn duetosquirmerrigidityareobtainedfromtheusualconstraintonthelocalrate-of-strain,namely
En
=
0foralln.SquirmerpositionsYn andorientationspn arethenupdatedwiththeLagrangianequations dYndt
=
Vn,
(27)dpn
dt
= Ä
n×
pn.
(28)InSection4,weshowthroughacomparisonwiththeboundaryelementsimulations from[28] thatourFCMsquirmer modelrecovers the velocities, angularvelocities, stresslets (Sn) andtrajectories fortwo interactingsquirmers for a wide
Fig. 2.ScalingoftheFCMwiththenumberofsquirmersNp.Computationaltimepertime-stepversusNp,orequivalently,versusvolumetricfractionφv.
Nc=256 coresworkinparallelforacubicdomainwith3843gridpoints.
2.3.1. Includingadditionalfeatures
WithFCM, additional effectscan readilybe incorporatedinto thesquirmer modelandinour subsequentsimulations, we considerseveralofthemtodemonstratetheversatilityofourapproach.Forexample,forcesduetostericinteractions andexternaltorquesexperiencedbymagnetotacticorgyrotacticorganismscanbeconsideredbyincludingtheminFn and
τ
n in Eq.(18). Additionally, we can extend the FCM squirmer model to ellipsoidal shapesby usingthe ellipsoidal FCMGaussian distributions.Such amodel canbe carefullytunedby comparingwithresultsfrom[55] and[56].The effectsof particle aspect ratioonsuspension properties canthen be explored systematically while still accountingforparticlesize effectssuchasJefferyorbits.Finally,wearenotlimitedtoconstantvaluesfor B1 andB2.Byallowingtheseparametersto
befunctionsoftime,theFCMsquirmermodelcanbeusedtoexplorehowtheswimmers’strokesaffectoverallsuspension dynamics.TheseextensionsareaddressedinSection6.
3. Numericalmethods
3.1. Fluidsolver
The smoothness of the Gaussian force distributions allows FCM to be used with a variety of numerical methods to discretizetheStokesequations.Ithasbeenimplementedwithspectralandspectral elementmethods[42,50,49]andfinite volumemethods[57,58]inbothsimpleandcomplexdomaingeometries.
Here,weuseaFourierspectralmethodwithFastFourierTransforms(FFTs) tosolvetheStokes equations(18)–(19),for thefluidflowinathree-dimensionalperiodicdomain.WesetthezerothFouriermodeofthevelocitytozero,u
ˆ
(
k=
0)
=
0, toensurenomeanfluidflowthroughtheunitcell.Asthefluidencompassestheentiredomain,thisisequivalenttoadding a uniform pressuregradient to balance thenet force the particles exerton the fluid inorder to maintainthesystem in equilibrium[59,37].Thisalsoensureswe haveaconvergentevaluationofthestressletinteractions[60].As thesquirmers can movewithoutexertinga forceonthefluid, wedopermit anetmaterialfluxofsquirmers throughtheunit cell.This correspondsdirectlytothefar-fieldStokesianDynamicscomputationsperformedin[30] forsquirmersuspensions.TheFFTs are parallelized withtheMPI libraryP3DFFT.This libraryuses2D decomposition ofthe 3D domain,introducing abetter scalability than FFT libraries that implementa 1D decomposition. This decomposition hasshown good scalability up to Nc=
32,
768 coresinDirectNumericalSimulation(DNS)ofturbulence[61].3.2. Computationalwork
Asexplainedin[49],thenumberoffloating pointoperationsforFCMscaleslinearlywiththenumberofparticles,Np.
WefindthesamescalingforourimplementationofFCM.Fig. 2showsthecomputationaltimepertimes-stepforNp upto
80,000particleswith3843
∼
6·
107 gridpointsandNc=
256 cores.3.3. Stericinteractions
IncludingstericrepulsionisstraightforwardwithFCM.Theseforcesareintroducedtobothpreventparticlesfrom over-lapping during the finite time-step and to account for contact forces. For spherical particles, we use the steric barrier described in[62].Forparticlesn andm, letrnm
=
Ym−
Yn andrnm= k
rnmk
.Therepulsive forceexperienced byn due toFbn
=
−
F2aref"
R2ref−
r2nm R2ref−
4a2#
2γ rnm,
for rnm<
Rref,
0,
otherwise. (29)where Fref is the magnitude of the force, the cut-off distance Rref sets the distance over which the force acts, and the exponent
γ
can be adjusted to control the stiffness ofthe force. Unless specified, all the simulations are run with Fref/
6π η
aU=
4, Rref=
2.
2a andγ
=
2.From Newton’s third law, we obtainFbm
= −
Fbn.It isimportant tonote thatthisforcebarrierisgenericanditisnotintendedtomodelanyparticularphysics.Whilewecouldinsteadusean exponential DLVO-likepotential[58],theforcebarriercanbeevaluatedmorerapidlythananexponentialpotentialandbyadjustingits parameters wecan incorporaterepulsionofa similar strengthandlength scaleasDLVO forces.Athorough studyonthe effectofthisforcebarrieronthedynamicsofparticulatesuspensionsisprovidedin[62].Stericrepulsionforspheroidal par-ticlesrequiresdifferentmodeling.Inourstudy,stericforcesandtorquesare introducedbyusingasoftrepulsive potential withthesurface-to-surfacedistanceapproximatedbytheBerne–Pechukasrangeparameter[63].
Usingadirectpairwisecalculation,theevaluationofstericinteractionsbetweenallparticlepairsateachtimestepwould requireO
(
N2p/(
2Nc))
computationspercore.ThiscostismuchgreaterthantheO(
Np)
costofthehydrodynamicaspectsofFCM.Therefore,insteadofadirectcalculation,weusethelinked-listalgorithmdescribedin[64].Thismethoddividesthe computationaldomainintosmallersub-domainsintowhichtheparticlesaresorted.Theedge-lengthofeachsub-domainis slightlylargerthan Rref.Thesesub-domainsaredistributedoverthecoreswherethestericinteractionsareevaluated.Fora homogeneoussuspension,thisresultsinaper corecost forsteric interactionsthat is O
¡
14N2p/(
2NcNs)
¢
,whereNs isthetotalnumberofsub-domains.Sinceforlargesystemsizes,wehaveNs
≈
L3/
R3ref≫
14 andNc≫
1,thelinked-listalgorithmforstericinteractionsismuchmoreefficientthanthedirectcomputation. 3.4.Algorithm
Wesummarizetheoverallproceduretosimulatelargepopulationsofmicro-swimmers inStokesflowwiththeFCM:
•
InitializeparticlepositionsYn(0)andorientationsp(n0),•
Starttimeloop:for k=
1,
. . . ,
Nit1. ComputeGaussians
1
n(k)(
x)
and2
n(k)(
x)
,Eq.(2),2. UpdateswimmingmultipolesG(nk),Eq.(13),andHn(k),Eq.(16),whichbothdependonpn(k),
3. Computestericinteractions,Eq.(29),withthelinked-listalgorithm, 4. Addadditionalforcingifany(gyrotactictorques,magneticdipoles,. . . ), 5. ProjecttheGaussiandistributionsontothegrid(RHSofEq.(19)),
6. SolveStokesequations,Eqs.(18)–(19),toobtainthefluidvelocityfieldu(k)
(
x)
,7. ComputeparticlerateofstrainsE(nk),Eq.(26),
8. If
k
En(k)k
>
ε
,computestressletsS(nk)following[49],a) Projectallthemultipolesontothegrid(RHSofEq.(19)),
b) SolveforStokesequations,Eqs.(18)–(19),toobtainthefluidvelocityfieldu(k)
(
x)
,9. ComputeparticlevelocitiesV(nk),Eq.(24),androtations
Ä
(nk),Eq.(25),10. IntegrateEqs.(27)and(28)usingthefourth-orderAdams–BashforthschemetoobtainYn(k+1) andp(nk+1).
4. Validation
4.1. ComparisonwithBlake’ssolution
AsexplainedinSection2.3,inFCMthevelocityfieldaroundanisolatedsquirmern inaninfinite,quiescentfluidisgiven by
uFCM
=
A·
Hn+
R:
Gn,
(30)whereAisasecondranktensorgivenbyEq.(21)andthethirdranktensorRcanbefoundin[38].Fig. 3shows Blake’s so-lutionEq.(12)and thevelocityfieldprovidedbyEq.(30)withtwodifferentvaluesforthedegeneratequadrupoleGaussian envelopesize,
σ
2 andσ
2/
2,thatappearsinthetensorA.Thestreamlinesareidentical,exceptfora near-fieldrecirculat-ingregion thatappears whenthewidthofthedegeneratequadrupole envelopeis
σ
2 (Fig. 3(b)).Inthisregion,however,themagnitudeofthevelocity issmallcompared tothe swimmingspeed. Thisregion shouldnot significantlyimpact the squirmer–squirmerhydrodynamicinteractionsthatweareaimingtoresolve.
Aquantitative comparisonofthevelocityfield isprovided inFig. 4.The agreementwithBlake’ssolutionis verygood forr
/
a>
1.
25 when usingσ
2 forthe width ofthe degenerate quadrupole envelope.As shown inFig. 4(b),the smallerenvelopesize (
σ
2/
2) matches Blake’ssolution moreclosely forr/
a<
1.
2, withclearimprovement atthe front andrearofthesquirmer.Forthisenvelopesize,thevelocity fieldinducedby thedegeneratequadrupoleuFCMB
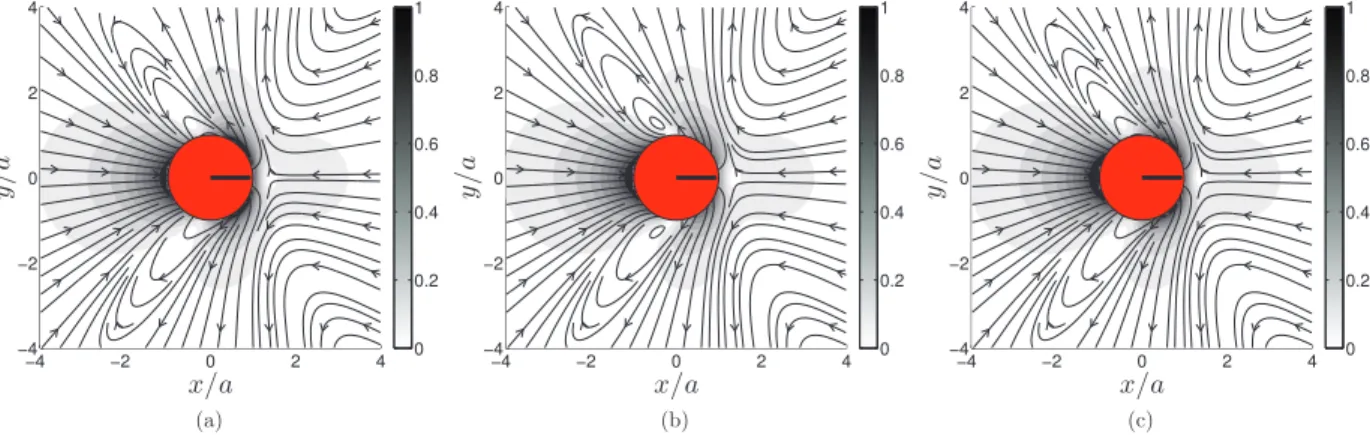
Fig. 3.Velocityfieldu/U aroundapullersquirmer(β=1)swimmingtotheright.(a)Blake’ssolution;(b)FCMsolutionwith σ2 forthedegenerate
quadrupoleenvelope.(c)FCMsolutionwithσ2/2 forthedegeneratequadrupoleenvelope.
Fig. 4.ComparisonwithBlake’ssolution,Eq.(12).(a)Normalizeddifference,kuBlake−uFCMk/U ,betweenFCMandBlake’ssolutionforapullersquirmer
(β=1).Thehalf-widthforthedegeneratequadrupoleenvelopeisσ2. :10% iso-value.(b)VelocityprofilealongtheswimmeraxisforBlake’s
solutionandtheFCMapproximationforthetwodifferentdegeneratequadrupoleenvelopesizes.
analytical solution uB1 in Eq.(12) (not shownhere). Below this width no quantitative improvement is observed as the
remainingerrorcomesfromthedipolarcontribution,uB2.
Whilethisquantitativecomparisonprovidesanicewaytochoosethedegeneratequadrupoleenvelopesize,wemustalso keep inmindthecomputational costassociatedwithdecreasingthislengthscale.Eventhough itwouldyieldaflowfield slightlymoreinregisterwithBlake’ssolution,resolvingthelengthscale
σ
2/
2 ina3Dsimulationwouldrequireagridwith8timesasmanypoints asthatneededfor
σ
2,thesmallestlength-scalealreadyinFCM.Thiswouldincreasecomputationtimesby atleastan orderofmagnitude. Inaddition,theFCM volume averagingusedto determinethesquirmer transla-tional andangularvelocities,Eqs. (3)–(5),will reduce thecontributionof thelocalizedvelocity field discrepancies tothe squirmer–squirmerinteractions.Inoursubsequentsimulations,wethereforeutilize
σ
2 forthedegeneratequadrupoleen-velopesizesincereducingthislength-scalewouldsignificantlyincreasethecomputationalcost,butonlyprovideaminimal improvement.
4.2. Pairwiseinteractionsofsquirmers
In[28],theauthorsperformeda varietyofsimulationsusingtheBoundaryElementMethod(BEM)tocomputetohigh accuracythepairwiseinteractionsbetweenasquirmerandaninertsphere,andbetweentwosquirmers.Here,weconsider thesamescenariosasthoseauthorsandcompareresultsfromourFCMsimulationswiththeirBEMresults.
4.2.1. Interactionsbetweenasquirmerandaninertsphere
Wefirstconsidertheinteractionsbetweenaninertsphere(labeled “2”)locatedatapointrfromthecenterofapuller squirmer(labeled “1”)with
β
=
5.Thedirectionr/
r formstheangleθ
withtheswimmingdirectionpofthesquirmer.The problemsetupisdepictedinFig. 5.Fig. 5.Sketch showing the set-up of our computations of the interactions between squirmer “1” and inert sphere “2”.
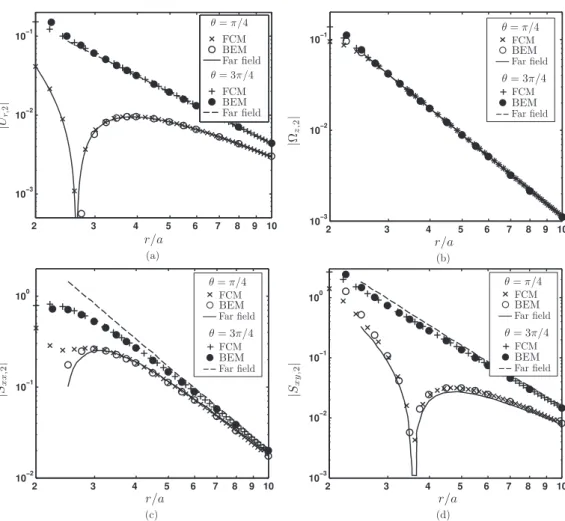
Fig. 6.The(a)radialvelocity|Ur,2|,(b)angularvelocity|Äz,2|,(c)stressletcomponent|Sxx,2|,and(d)stressletcomponent|Sxy,2|fortheinertsphere“2”
atadistancer fromapullersquirmer(β=5).
Fig. 6comparesthevelocityofthesphereobtainedusingFCMsimulationswiththeBEMresultsandfar-fieldanalytical solutionsfrom[28].Wecomputedthefar-fieldvelocityinFig. 6(a)usingtheexpressionsprovidedin[28] sinceitwasnot plottedin Fig. 6(a) of [28] forr
/
a<
2.
5. We suspect that the BEM resultswere also not plottedfor thisrange.As FCM alsoresolves themutuallyinduced particlestresslets,itprovides amoreaccurate estimationthan far-fieldapproximation of[28].Asaresult, weseethattheFCM resultsverycloselymatchBEM,evenintherangewherer/
a<
3.Similartrends areobserved forthe angularvelocity oftheinertsphere (Fig. 6(b)) andthestressletcomponents (Figs. 6(c), 6(d)).These comparisonsillustratetheaccuracyoftheresultsthatcanbeobtainedusingFCM,whichcloselymatchesBEMbutincursa fractionofthecomputationalcost.Fig. 7.Initial configuration of the squirmers in the trajectory simulations.
Fig. 8.Trajectoriesoftwosquirmersswimminginoppositedirectionswithtransverseinitialdistanceδy=1a,. . . ,10a.Lines:datafrom[28].Symbols:FCM results.
4.2.2. Trajectoriesoftwointeractingsquirmers
Wecomputethetrajectoriesoftwopullersquirmers(
β
=
5)andcomparetheresultswiththeBEMsimulationsof[28]. One squirmerinitially swimsin thex-direction, p1=
ex,andtheother one intheopposite direction, p2= −
ex. Theyareplaced with initial separation distance
δ
y=
1a,
. . . ,
10a inthe transverse direction andδ
x=
10a in the x-direction. The problemset-up isdepictedinFig. 7.Since thesquirmers maycollide,we alsoincludesteric interactionsprovided bythe forcebarrier(29).As shownin Fig. 8, the trajectories match very well with the BEM results for
δ
y≥
2a. Whenδ
y=
1a, the collision barrier andnear-field hydrodynamic interactions play an importantrole in determiningthe overall squirmer trajectories.Fig. 9showstheeffectofthestericrepulsionparameters Fref andRref on thesquirmertrajectories.The specificvaluesof theseparameters are provided inAppendix B.We seethat by varying thebarrier parameters onecan obtain trajectories thatcloselymatchtheresultsof[28].
5. Simulationresults
5.1. Largesuspensionsofswimmingmicro-organisms
The squirmer model has been widely used to investigate both the behavior of singleswimmers [65–68], aswell as their collectivedynamicsandinteractions [29,30,69,34,33].Simulationsofsuspensionsrevealedthattheoverallpopulation dynamicsdependstronglyonthesquirmingparameter
β
.Inparticular,when|β|
issmall,theisotropicstateforaperiodic suspensionhasbeenshowntobeunstable,andthesuspensionevolvestoapolarsteady-statewithanon-zerovalueofthe polarorderparameterP
(
t)
=
¯
¯
¯
¯
¯
¯
1 Np NpX
n=1 pn(
t)
¯
¯
¯
¯
¯
¯
.
(31)Fig. 9.Influenceofthecollisionbarrierparametersonthesquirmertrajectoriesforthecasewhereinitiallyδy=1a.(a)Trajectories.Crosses:datafrom
[28].(b)Forcebarrierprofiles(seealsoAppendix B).Thelinestylescorrespondthoseshowingthetrajectoriesin(a). :barrierusedinFig. 8.
Fig. 10.Snapshotsoftheorientationalstateinasemi-dilutesuspension(φv=0.1)containingNp=37,659 swimmerswithβ=1.hpiisthemean
steady-stateorientationvectorontheunitspheredefinedinEq.(32).pn· hpiquantifiesthedegreeofalignmentwiththemeandirectionhpiofswimmer n.
Thisinstabilityhasbeenstudiednumericallyby[30] and[69] usingStokesian dynamicswithNp
=
64 swimmersforvol-umefractions
φ
v=
0.
01→
0.
5.UsingtheLattice-Boltzmannmethod,[34]observedthesamebehavior for Np=
2000 andφ
v=
0.
1.5.1.1. Polarorderparameter
Using ourFCM model, we studythisinstability andthe resulting polarorderof a squirmersuspension. In particular, we examinehowthe domainsize affectsboth thegrowthrateofthe instabilityandthe finalsteady-state.The influence ofdomain size hasnot been addressedpreviously for squirmer suspensionsthough it hasbeen observed insimulations ofrod-likeswimmers [25]. We performsimulations of semi-dilutesuspensions (
φ
v=
0.
1) of pullersquirmers (β
=
1) intriply-periodicsquaredomains withedgelengthsrangingfromL
/
a=
14 to L/
a=
116.Asthevolume fractionφ
v isfixed,varying L
/
a increasesofthenumberofswimmersfrom Np=
64 (asin[30,69]) to Np=
37,
659.Weinitializeahomoge-neous,isotropicsuspensionby distributingtheswimmer positionsuniformlyinthedomainandtheswimmingdirections uniformlyovertheunitsphere.Dependingondomainsize,thesimulationsareruntofinaltimetf
=
1000−
1500a/
U withatimestep
1
t=
0.
005a/
U .Thus,eachsimulationrequiresbetween2×
105–3×
105time-steps.Fig. 10illustrates the polar ordered state for a simulations with L
/
a=
116, Np=
37,
659. We quantifythe degree ofalignmentofeachswimmern,pn
· h
pi
,withthemeansteady-stateorientationh
pi
givenbyh
pi =
lim t→∞ 1 Np NpX
n=1 pn(
t)
P(
t)
,
(32)where
h
pi
lies on the unit sphereS
2.At t=
0, there is no clear mean orientation, while at t=
1000a/
U ,a significant proportionofparticlesarealignedwiththemeandirectionh
pi
.Fig. 11.PolarorderP(t)inasemi-dilutesuspension(φv=0.1)ofsquirmerpullers(β=1).(a)Timeevolutionofpolarorderdependingonthenumberof
swimmers. :L/a=14,Np=64; :L/a=19, Np=174; :L/a=38,Np=1395; :L/a=58,Np=4707; :L/a=77,
Np=11,158; :L/a=116,Np=37,659.(b)SteadystatevalueP∞dependingonNp∼ (L/a)3. :fitlinearwith1/pNp.(Inset):Dependence
ofP∞with1/pNp.
Fig. 12.Characterizationofthepolarinstability.(Mainfigure):Timeevolutionofpolarorderinsemilogarithmicscalesuggestsanexponentialgrowthof theinstability.(Inset):GrowthrateoftheinstabilityfordifferentNp(andL/a).
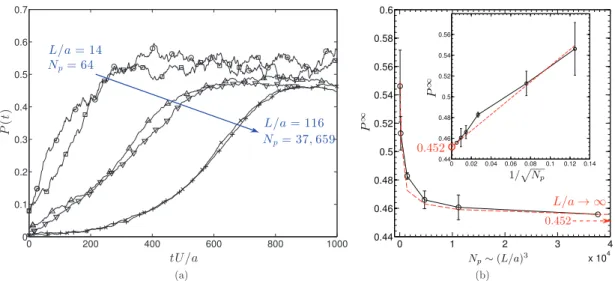
Fig. 11ashows P
(
t)
,Eq.(31),for thesimulations with differentdomain sizes.For each domainsize, we see that the suspension evolvesfromtheinitial isotropicstate to onethat haspolarorder. Weobserve,however,that the finalvalue, P∞,ofthepolarorderparameter dependsonthedomain size.We findthatit decreases asL/
a (and Np) increases.ThedataalsoshowsthatasL
/
a→ ∞
, P∞ decayslike(
L/
a)
−3/2 (or N−1/2p )andreachesanasymptoticvalueof P∞
→
0.
452.Theseresultswouldindicate thatpolarordershould alsoariseinan unbounded suspension.Wealso observethatinthe polarorderedstate, theaverageswimmingspeed is4%less thanitsvalue inisotropicstatewhichitselfisnearly thefree swimmingvalue.Indeed,weremindthatinoursimulations,azeronetfluxofthesuspensionisimposed,therefore,when theswimmersarealigned,theyaresloweddownbythebackflow.
Fromoursimulations,wemayalsoanalyze howL
/
a affectsthetimeevolutionoftheinstability.InFig. 11(a),weclearly seethatthetimeittakestoreachthefinalpolarstate increaseswithdomainsize.WeanalyzethisdatainmoredetailinFig. 12,nowplottingitin semilogarithmicscale. Foreach case,we findthat afteraninitial transientstate, theinstability growsexponentiallyandwithagrowthratethatisindependentofthesystemsize(seeinsetfigure).Itwouldcertainlybe interestingtoinvestigateifthisresultcouldbereproducedbyalinearstabilityanalysisofcontinuummodelforasquirmer suspension.
5.1.2. Orientationaldistribution
Wecan examinethepolarorderinmoredetailbycomputingtheorientationaldistribution
9(θ,
φ,
t)
definedoverthe unitsphere[70].Here,θ
=
cos−1(
pz)
correspondstotheelevationanglewhileφ
=
tan−1(
py/
px)
givestheazimuthalangle.Fig. 13.Orientationaldistributionontheunitsphere(φ,θ ).(a)Distributionbeforethetransitiontopolarorder:tU/a=90.(b)Distributiononcethepolar orderedstateisreached:tU/a=1110.
Fig. 14.Time-averagedsteady-stateorientationaldistributionintheframeofthemeanorientationvector9(θ,φ)|hpi.(a)Distributionovertheunitsphere.
(b) :distributionofelevationangleθaveragedoverazimuthalangleφ; :uniformdistribution90(θ )=1/π; :distributionof
az-imuthal angleφaveragedoverelevationangleθ; :uniformdistribution90(φ)=1/(2π).
Fig. 13shows
9(θ,
φ,
t)
normalizedby theisotropicdistribution90
=
1/(
4π
)
attimesbeforeandafter thetransitionto polarorder for the case where Np=
11,
158 and L=
78a. As expected, before the transition, the distribution is nearlyuniformoverthesurfaceofthesphere.Afterthepolarstateisreached,weseethat
9(θ,
φ)
isnarrowlydistributedaround the mean directionh
pi
. We note that the steady state mean direction depends both on the random initial seeding of swimmers and the domain geometry. We find that system first aligns along an arbitrary direction due to the random initialseedingofswimmers,butastimegoeson,themeandirectortendstoalignwiththenormaltooneoftheperiodic boundaries.Asaconsequence,thesteadystatemeandirectionisbiased bythedomainshapethatbreaksradialsymmetry. We would like to stress that, however, asdemonstrated here andin [30], the polarordering itself is a consequence of squirmer hydrodynamic and steric interactions rather than the domain shape. Fig. 14 shows the steady-state averaged distribution9(θ,
φ)
|
hpi in the frame where the mean direction isgiven byθ
=
0 andφ
=
0. We see that the resultingdistributionisaxisymmetricinthatitdoesnotdependon
φ
.Thisisnotsurprisingastheflowfieldinducedbyasquirmer isaxisymmetric.Orientationaldistributionscouldalsobe obtainedfromcontinuummodels,aswasdoneforswimmersin externalflow fields[70]. Ourresultscouldbe comparedwithcontinuum models ofsquirmersuspensions,though,to the bestofourknowledge,thecontinuum modelsthat arecurrentlyavailable intheliterature predictastableisotropicstate forsphericalswimmersuspensions.5.1.3. Spatialdistribution
Alongwiththeorientationaldistribution,wealsoexaminethespatialdistributionofsquirmersbycomputingtheVoronoi tessellation forthe set of points corresponding to the squirmers’ centers. We have used the C++ libraryVoro++ [71] to
Fig. 15.DistributionofVoronoicellvolumes.(Mainfigure):Timedependenceofthestandarddeviation,σV,oftheVoronoivolumedistributionnormalized
bythemeanvaluehVVi.(Inset):TimeaverageoftheVoronoivolumedistributionbefore( )andafter( )thepolarordertransition.
Fig. 16.Steady state pair distribution function (a) g(r, θ ). (b) g(r).
performthecomputationanddeterminethevolume, VV,ofeachVoronoicell.Fig. 15showstheevolutionofthestandard
deviation,
σ
V, of the VV distribution for the case where Np=
11,
158 and L=
78a. We see sudden increase inσ
V asthe suspensiontransitions topolarorder,corresponding toa wideningofthedistribution.The insetinFig. 15 showsthe time-averaged distribution before and after the transition. Before polarordering occurs, the mode ofthe distribution is
¯
VV
≈
34a3,which is slightlyless than theaverage valueh
VVi
=
L3/
Np=
42a3 that one might expect.During the polarsteady-state,wefindthatthemodedecreasesto V
¯
V≈
30a3,whichisindicativeofaslightclusteringoftheparticles.Tofurtherexaminethespatialdistribution,wecomputethesteady-statepairdistributionfunctiong
(
r,
θ )
whichgivesthe probabilityoffindingasquirmeratadistancer= |
r|
andwithelevationangleθ
=
cos−1(
r·
p/
r)
fromanothersquirmerthat hasswimmingdirectionp.Fig. 16(a)showsg(
r,
θ )
forthecasewhereNp=
11,
158 andL=
78a.Weseeclearlythatg(
r,
θ )
dependsnotonlyonr,buton
θ
aswell. Atsteady-state,foragivensquirmerthereisasignificantly higherprobability of finding anothersquirmer infront ofit (θ
=
0) ratherthan behind it.If we integrate g(
r,
θ )
overθ
,we obtain theradial distribution functionshownin Fig. 16(b).Wefind that g(
r)
exhibitsa peakattwo radii fromtheswimmer surface. This peaksuggeststheexistenceofparticleclusterswhosesizesaregreaterthantwoindividuals.5.1.4. Orientationalcorrelations
The hydrodynamic and steric interactions between the squirmers also lead to correlations in their orientations. We computethesteady-stateorientationalcorrelationfunction[30]
Fig. 17.Correlations in squirmer orientation at steady state. (a) Ip(r, θ ). (b) Ip(r).
where the brackets
h·i
denote the ensemble average that we compute by averaging over both time andsquirmer pairs.Fig. 17(a)showsthecorrelationsinthe frameofasquirmer locatedattheoriginandwithp
= ˆ
z andthe definitionsofr andθ
areidenticaltothoseinSection 5.1.3.Wefindthat thehighestcorrelationsoccurnearcontact(r≈
2)attheanglesθ
≈
π
/
3 andθ
≈
3π
/
4.Despitetheoverallpolarorder,wecanidentifytwodistinctregionsaroundθ
=
0 andθ
=
π
where theorientationsareuncorrelated.Wedo,however,findapositivevalueforthecorrelations,evenfarawayfromtheorigin. Thisismostevidentintheθ
-averagedcorrelation, Ip(
r)
,showninFig. 17(b)whichapproachesafinitevalue Ip=
0.
22 asr increases.Thisisconsistent withtheobservedlong-rangepolarorderofthesuspension.We alsofindthat Ip
(
r) >
0 forall r.Thisagainmaybearesultofthestrongpolarorderingofthesuspension. 6. Extensionstoellipsoidalswimmersandtime-dependentswimminggaits
Inthissection,wedemonstratetheextensionofourapproachtobothspheroidalswimmershapesandtime-dependent swimming gaits. This illustrates the versatility of FCM while preserving good computational scalability and an accurate treatmentofhydrodynamicinteractions.
6.1. FCMforellipsoidalparticles
ToextendFCMtopassiveellipsoidalparticles,onesimplymodifiestheGaussianenvelopesinEq.(2) [50].Forexample, takingthe orthonormal vectors e
ˆ
1, eˆ
2, and eˆ
3 to be aligned withthe ellipsoid semi-axes having lengths a1, a2, and a3respectively,theGaussianenvelopecorrespondingto
1
n(
x)
is1
elln(
x)
= (
2π
)
−3/2(
σ
1;1σ1;2σ1;3)
−1exp·
−
12(
x−
Yn)
TQ
T6
1Q
(
x−
Yn)
¸
(34)
where
σ
1;i=
ai/
√
π
fori=
1,
2,
3,Q
= (ˆ
e1 eˆ
2 eˆ
3)
,and6
1=
σ
1−;21 0 0 0σ
1−;22 0 0 0σ
1−;23
.
(35)Asimilarexpression isusedfor
2
elln(
x)
withσ
2;i=
ai/
¡
6
√
π
¢1
/3.Beyondthis,theunderlying algorithmofprojectingthe particleforcesontothefluidandvolumeaveraging theresultingfluidflowremains unchanged.Theconstraintthat En=
0foreach n isstill usedto find thestresslets. Thus, usingFCM to compute the motionand hydrodynamic interactions of ellipsoidsdoesnotrequireanyadditionalsteps inthealgorithmdescribed inSection 3.4.AsexplainedinSection 3.3,the modeling ofstericinteractionsdiffers fromthat forsphericalparticles andoursimpleforce barriercouldnotbe used.An extensivevalidationofFCMforellipsoidalparticlesispresentedin[50] wheremanyoftheclassical resultsforellipsoidal particles inStokes flow, e.g. Jeffery’s orbits,are shownto be recovered exactlywith FCM.We note that for veryslender particles,FCMwouldnotbeappropriateandcomputationsbasedonslender-bodytheories[72]shouldbeperformed.
Fig. 18.Simulationofadilutesuspensions, φv=0.05,ofprolate spheroidalpushers,B2= −1.5, B1=0,withaspectratio aa12 =
a1
a3 =3.(a)Snapshot
att=30U/a1.Colorsindicatethevelocitymagnitudenormalizedbytheindividualswimmingspeed.(b)TimeevolutionofpolarorderP(t). :
isotropicvalueofthepolarorderparameter1/pNp=0.026.
To extend FCM to active ellipsoidalparticles requires addingthe stresslet and possible potential dipole termsto the multipoleexpansion.Asforsphericalparticles(seeSection2.3,Eqs.(20)and(23)),theseadditionalmultipoleswillleadto artificial, self-inducedvelocitiesandlocalrates-of-strain.Theseeffectsmustbesubtractedaway usingtheformuladerived inAppendix A.ItisworthreiteratingthatthesearetheonlystepsthatneedtobeaddedtotheFCMalgorithmforpassive particlestosimulateactiveones.
6.2. Spheroidalswimmersimulations
Previousparticle-basedsimulations[25,32]andcontinuumtheoryresults[18–20]predictanunstableisotropicstatefor suspensionsofprolatespheroidalpushers.UsingFCM,wesimulateadilutesuspension,
φ
v=
0.
05,ofNp=
1500 spheroidalpushers with aspect ratio a1 a2
=
a1
a3
=
3. For these simulations, the stressletparameter is B2= −
1.
5 and the degeneratequadrupole isset tozero, B1
=
0. Stericinteractions are includedby usinga softrepulsive potential witha∼
r−5 decayandsurface-to-surfacedistanceapproximatedbytheBerne–Pechukasrangeparameter[63].Fig. 18(a)showsasnapshot of thesuspension whereclustersofswimmershavevelocities nearly1
.
6 timeslargerthan theisolatedswimmingspeed.As in[25,32],weseethattheisotropicstateisnot stableandthepolarorderparameterincreaseswithtime(Fig. 18(b)).We donotseeaslargean increasein P(
t)
asin[25,32]duetotherelativelylowaspectratioofourswimmers.Thetumbling effectduetoJefferyorbitsandthesterictorquesthat tendtoalignadjacentswimmersaremuchlower thanthey arefor rod-likeswimmers.6.3. Suspensiondynamicsoftime-dependentswimmers
Here,weshowhowtoincorporatetime-dependenceintoourmodel.Usingtheprocedureoutlinedin[73],wedetermine thetime-dependentmultipolecoefficients B1
(
t)
andB2(
t)
fromtheexperimental dataprovidedin[39].Wethenperform suspension simulations thatshow time-dependenceatthelevel ofindividualswimmerscan affecttheoverall suspension properties.6.3.1. Asingletime-dependentswimmer
Recentexperiments[39]quantifiedtheperiodicswimminggaitandresultingflowfieldofthealgaecellChlamydomonas Rheinardtii.Theyextractedtheswimmingspeed,theinducedvelocityfield,andthepowerdissipation,showingalsothatall canberepresentedasperiodicfunctionsoftime.Arecenttheoreticalinvestigation[73]showedthatthesequantitiescould bereproducedusingamultipole-basedmodel.Intheirstudy,theyconsideredthreetime-dependentmultipoles:a stresslet, adegeneratequadrupole(orpotentialdipole)anda“septlet”.Thestressletdecaysasr−2whereasthedegeneratequadrupole
andthe“septlet”decayliker−3.Here,weutilizeonlythestressletanddegeneratequadrupoletermsandfindthattheyare
sufficienttoreproducethefeaturesmeasuredby[39].
Themeasuredswimmingspeedfrom[39],U
(
t)
,canberepresentedusingthetruncatedFourierseriesintime(see[73]):U
(
t)
=
a0+
a1cos(
ω
t)
+
a2cos(
2ω
t)
+
b1sin(
ω
t)
+
b2sin(
2ω
t)
(36)where
ω
is the frequencyof the swimming gait. The mean swimmingspeed over one beatperiod, T=
2π
/
ω
, is given by a0=
49.
54a s−1,where a=
2.
5 µm is the radius ofthe microorganism. The values forthe remaining coefficientsareFig. 19.(a) (B1(t), B2(t)) phase diagram for one beat cycle. (b) Power dissipation5d(t)over one beat cycle. : FCM, : results from[39].
providedinthe supplementaryinformationof[73].From Eq.(9),we canimmediatelydetermine thetime-dependent de-generatequadrupolestrength
B1
(
t)
=
3
2U
(
t)
(37)inordertopreserve theinstantaneousforce-freecondition. Unlikethisterm, thereismorethan oneway tocalibrate the stressletstrength,B2(t
)
.Forexample,onecoulddetermineB2(t)
usingthepowerdissipationmeasurementsfrom[39] and Eq.(9)from[74]5
d(
t)
=
2 3π η
a³
8B1(
t)
2+
4B2(
t)
2´
(38)forthe power dissipated by a squirmer. Using this approach,we found that ourresulting flow field did not match the experimental results from [39]. We instead determine B2(t
)
directly from the experimental flow field by fitting to the location of the moving stagnation point. Thisis the same approach adopted by [73]. We utilize three Fouriermodes to describethetimeevolutionof B2B2
(
t)
=
c0+
c1cos(
ω
t+
ϕ
c1)
+
c2cos(
2ω
t+
ϕ
c2)
+
c3cos(
3ω
t+
ϕ
c3)
+
s1sin(
ω
t+
ϕ
s1)
+
s2sin(
2ω
t+
ϕ
s2)
+
s3sin(
3ω
t+
ϕ
s3) ,
(39)wherethe amplitudes c0,c1,s1,
. . .
andphasesϕ
c1,
ϕ
s1,
. . .
are fittedmanually. Appendix C givesthe resulting valuesoftheseparameters.
ThephasediagraminFig. 19(a)showsthevalueof B1 versus B2 anditissimilartothatfoundby[73].Weextractthe averagevalueof
β(
t)
=
B2(
t)/
B1(
t)
overonebeatcycle¯
β
=
1 T TZ
0 B2(
t)/
B1(
t)
dt=
0.
1 (40)whichcorresponds to apullersquirmer witharelatively smallstressletmagnitude. Fig. 19(b)showsthe resultingpower dissipationasdetermined fromEq.(38).Itreachesapeakvalueatt
/
T≈
0.
3,whichcoincideswiththetime atwhichthe swimmingspeed reachesits maximumvalue. We note that unlike [39] ourswimmers generateaxisymmetric flowfields andweareconsideringa3Dperiodicdomain.Despitethis,weachieveaqualitativelysimilarpowerdissipationprofilewith slightlygreatervaluesduringthefirsthalfofthebeatcycle.Fig. 20showstheflow fieldaround ourmodelofC.Rheinardtii atsixdifferenttimesduringits beatcycle.Thesetime points arechosen to correspond tothose inFig.3 of[39]. Weachieve very similar streamlinesand, by construction,the positionof thestagnation point matches theexperimental data very well. We alsonote that our flow field is similar to thatgivenbythemultipolemodelfoundin[73]eventhoughwedonotincludetherapidlydecaying“septlet”terminour model.Theseresultsillustratethatourproperlytuned,time-dependentsquirmermodelcanyieldflowfieldsverysimilarto thoseofrealorganisms.
Fig. 20.Snapshotsofthetime-dependentflowfieldaroundamodelChlamydomonas.Backgroundgreylevelsrepresentthenaturallogarithmofthenorm ofthevelocityfieldinrad s−1. :positionofthestagnationpointgivenbytheFCMmodel. :positionofthestagnationpointmeasuredby[39].(Insets)
Swimmingspeedalongthebeatcycle.
Fig. 21.RelativetrajectoriesoftwomodelC.Rheinardtii swimminginoppositedirectionswithstrokephaseshift1ϕandinitialtransversedistanceδy=1a and2a.Thegreylevelslightenas1ϕincreasesfrom1ϕ=0→ 1ϕ=7π/4. :trajectoryoftwosteadysquirmerswithβ=0. :trajectoryof twosteadysquirmerswithβ=0.1.(Top)δy=1a.(Bottom)δy=2a.
6.3.2. InteractionsbetweentwomodelC.Rheinardtii
We first consider pairwise interactions betweentwo model C.Rheinardtii for two initial configurations,
δ
y=
1a and 2a (cf. Section 4.2.2). We introduce a phase shift,1
ϕ
, between their swimming cycles. When1
ϕ
=
0, the swimmers are synchronized,whereas for1
ϕ
=
π
they are completelyout of phase.Fig. 21 showsthe effectofthisphase shifton the trajectories. For each case, we show only the trajectory of the swimmer labeled “2” in Fig. 7. We also provide the trajectories ofsteady swimmerswithβ
=
0 andβ
=
0.
1 for comparison. Recall thatβ
=
0.
1 corresponds to the average value of B2(
t)/
B1(
t)
in ourmodel.Wesee thatthe trajectories,includingthefinal positionsandorientations,dodependFig. 22.Sketchofthesimulationswithtime-dependentswimmers.Eachswimmern generatestime-dependentflowdisturbanceswiththesameamplitude andfrequency,butthephase1ϕnmaybedifferentforeachswimmer.
on
1
ϕ
.Wefind,however,thatthesevariations aresmallcomparedto theswimmerseparation distance(see theinsetofFig. 21fortherelativetrajectoriesintheframeofoneswimmer).Thetrajectoriesarealsoclosetothoseforsteadysquirmers with
β
=
0,
0.
1.6.3.3. Collectivedynamicsoftime-dependentswimmers
Inthissection,wepresentresultsfromsuspensionsimulations usingourunsteadymodelforC.Rheinardtii. Tothebest of our knowledge, similar results have not appeared in the literature. We aim to show in this initial study that time-dependenceofindividualswimmerscanaffecttheir overalldistribution.Specifically,weshow thatthedistributionofbeat phasesaffectsthesteady-statepolarorderofthesuspension.Fig. 22showsasketchofthesimulationswithtime-dependent swimmers.
Weconsider asuspension of Np
=
1395 time-dependentswimmers distributeduniformlyina triply periodicdomain.Thevolumefractionis
φ
v=
0.
1.Wetake theinitialswimmingdirectionstobedistributeduniformlyovertheunit-sphere.Forswimmern, itsgaitis characterizedby B1
(
t+ 1
ϕ
n)
and B2(
t+ 1
ϕ
n)
, where1
ϕ
n isthe valueof itsbeatphase. Weconsidertwodifferentdistributionsofthebeatphases.Thephasesareeitheruniformlydistributedwith
1
ϕ
n∈ [
0;
2π
]
,or1
ϕ
n=
0 for all n such that all swimmers are synchronized.We run the simulations for3700 dimensionlesstime units,corresponding to 4000beat cycles.As mentioned in Section 6.3.1, the meanvelocity of C.Rheinardtii is U
=
49.
54a s−1.Sincewetakethetime-step
1
t=
0.
0025a/
U ,thetotaltimeforoursimulationscorrespondto1.
5×
1061
t.Fig. 23(a)showstheevolutionofthepolarorderparameterforthesetwodistributionsofbeatphase.Alsoshownisthe polarorderparameterforsuspensionsofsteadyswimmerswithswimmingspeedsequaltothemeanswimmingspeedfor thetime-dependentcaseandeither
β
=
0 orβ
=
0.
1.Wefindthatwhenthereisauniformdistributionofthebeatphase, weachieveresultsthatmatchthesteadycasewithβ
=
0.
1.Ontheotherhand,iftheswimmersaresynchronized,wefind thatthepolarorderparametermatchesthatforthesteadycasewithβ
=
0.Fig. 23(b)showsthesteady-stateorientational distributions9(θ )
|
hpiaboutthemeandirector.Again,weseethatthedistributionforcaseofauniformdistributionofbeatphasesmatchesthatforthesteadycasewith