NRC Publications Archive Archives des publications du CNRC
Texte intégral
Figure
Documents relatifs
Le système classique pour l'évacuation des eaux pour les toitures plates ou à faible pente consiste d'orifices d'écoulement, installées sur la face du toit,
Here, we have shown that while all copper nanoparticles tested exhibit cytotoxicity against all three cell lines tested, particles in the 40–60 nm range appear to affect the cells
Bacteria grown in the presence of Neu5Ac or Kdn were incubated with 10% pooled normal human sera for 5 min at 37°C and IgM antibody (A) and complement factor C3 (B) deposition
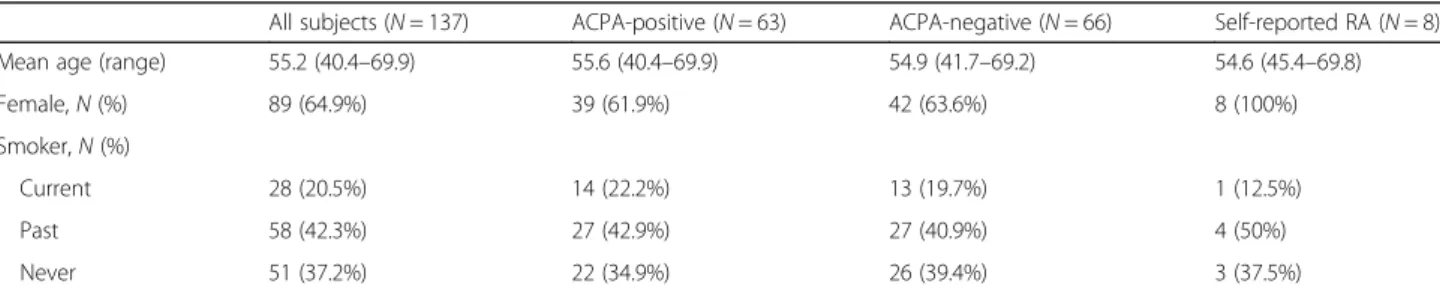
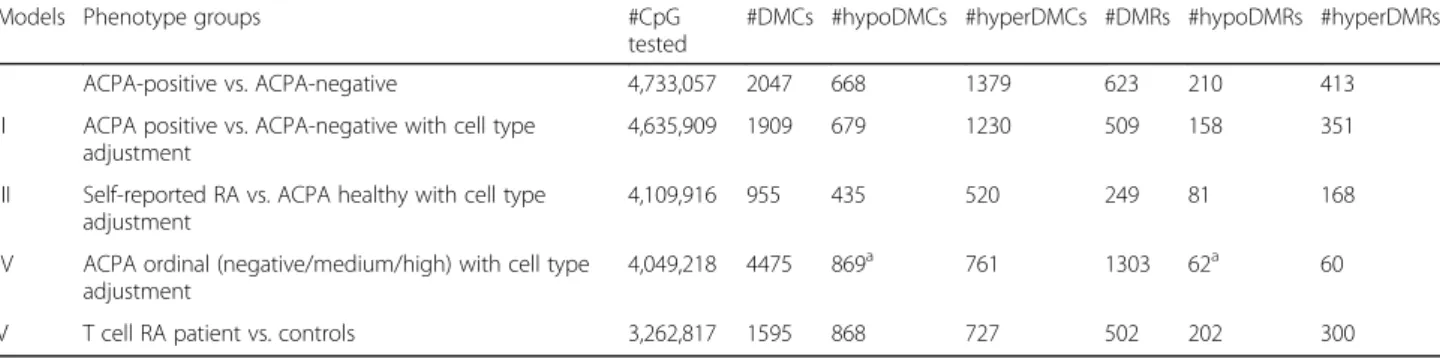
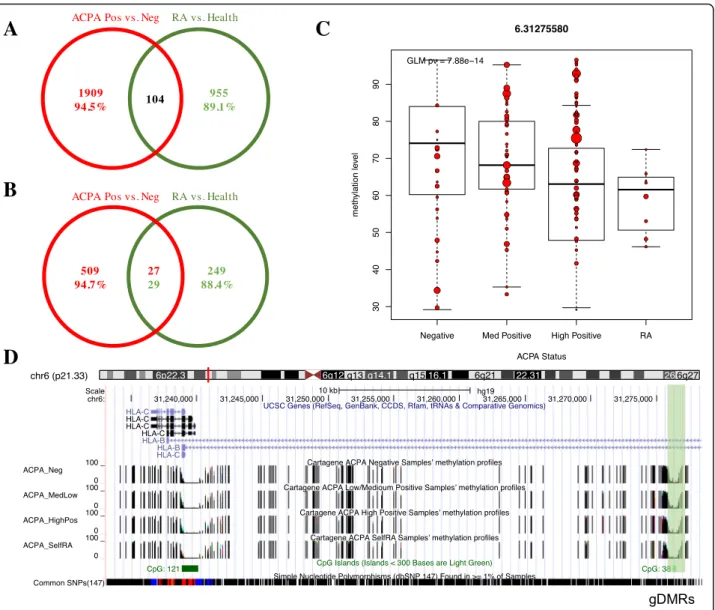
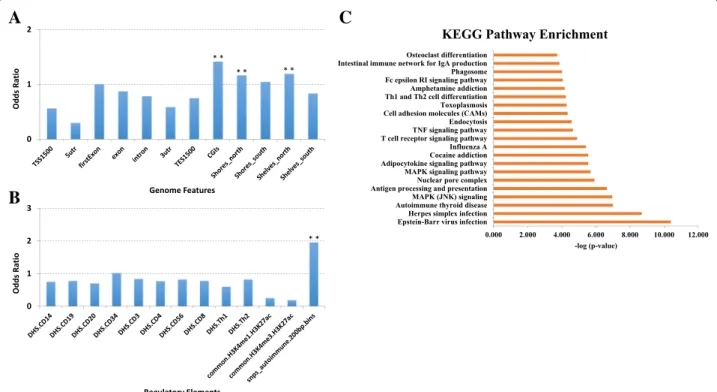
(C) TdDof gene expression levels, with greater expression in solid-stemmed lines W9262-260D3 and CDC Fortitude (blue bars), lower expression in hollow-stemmed lines Svevo and Kofa,
Seasonal habitat drives intestinal microbiome composition in anadromous Arctic char (Salvelinus alpinus).. Element, Geraint; Engel, Katja; Neufeld, Josh D.; Casselman, John M.; van
As the microspores from BnA9::BnaC.MAGL8.a plants coalesced with each other in stage 11, when the pollen coat substrates was still enclosed in tapetum, the sticky pollen phenotype
Our results are clear: a combination of practical operational modifications (AC cycling, doing laundry later in the evening) and commercially-available technology (exterior
Access and use of this website and the material on it are subject to the Terms and Conditions set forth at Diversity array technology markers: genetic diversity analyses and
