Microorganisms, flight, reproduction, and predation in birds
232
0
0
Texte intégral
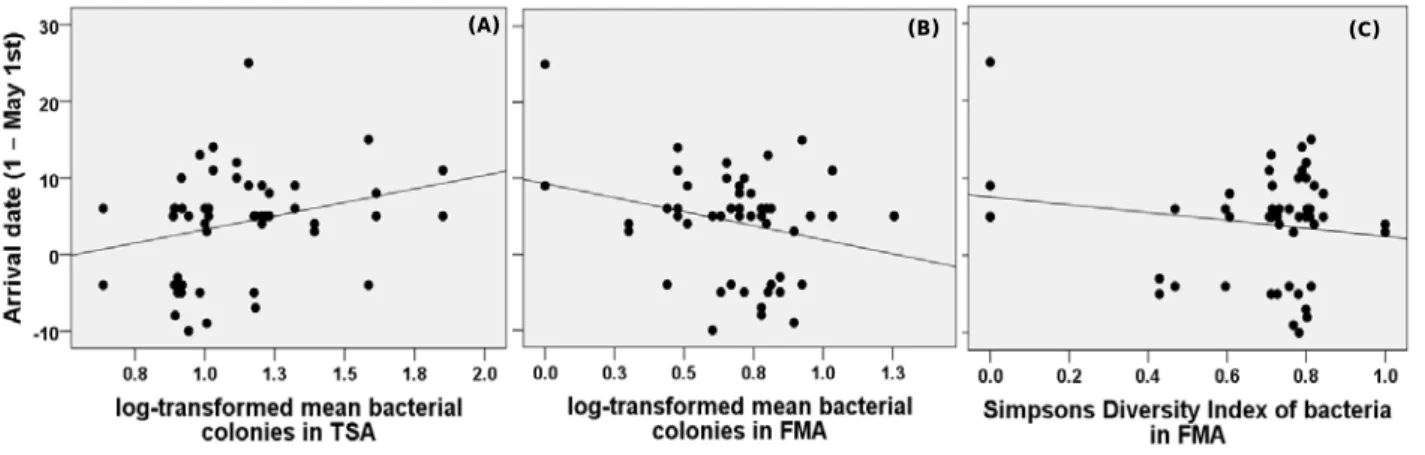
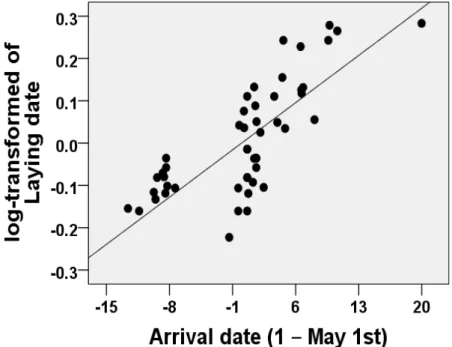
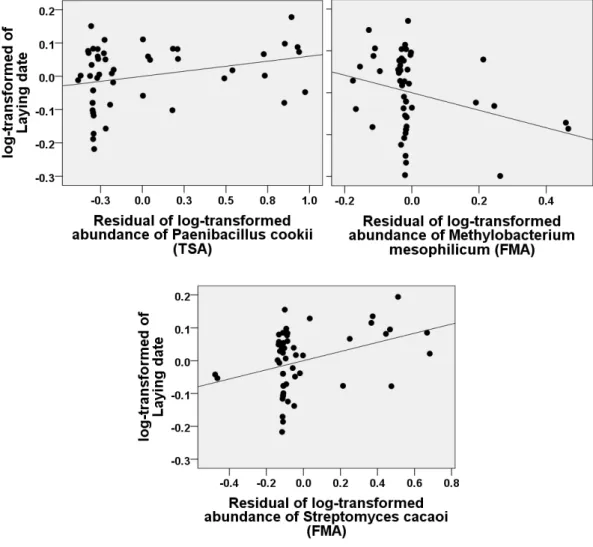
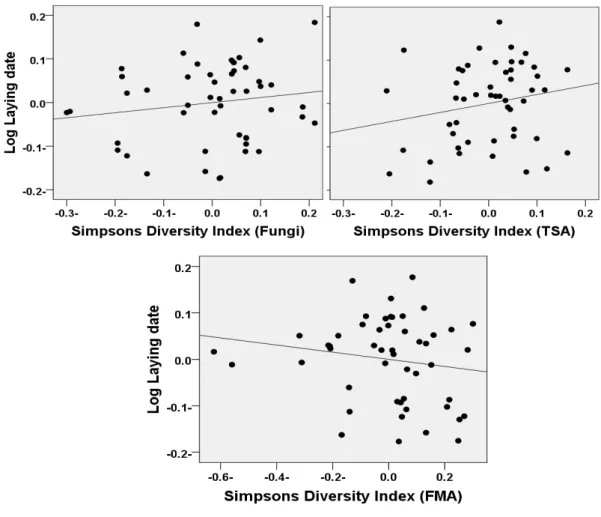
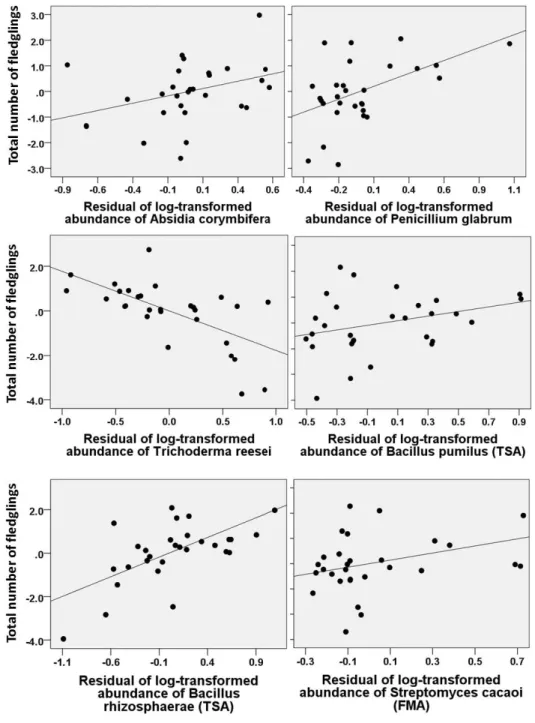
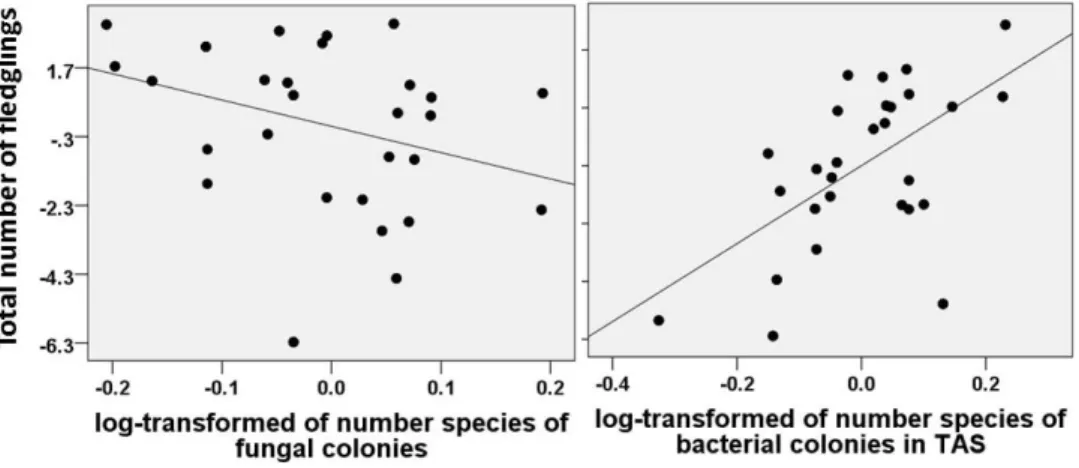
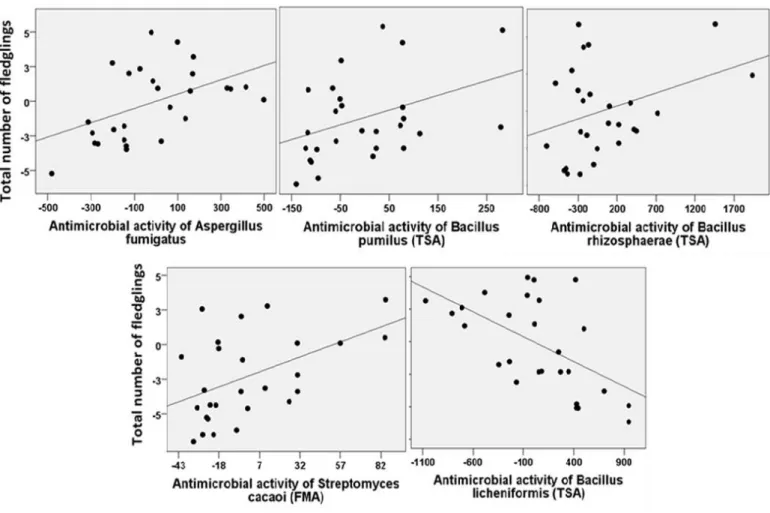
Figure



+7
Documents relatifs