Multiscale population dynamics in reproductive biology: singular perturbation reduction in deterministic and stochastic models
Texte intégral
Figure

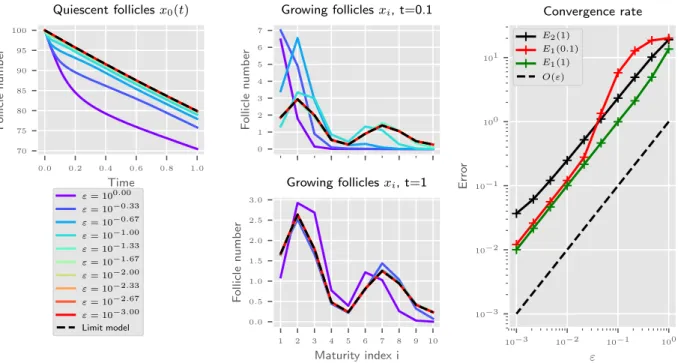


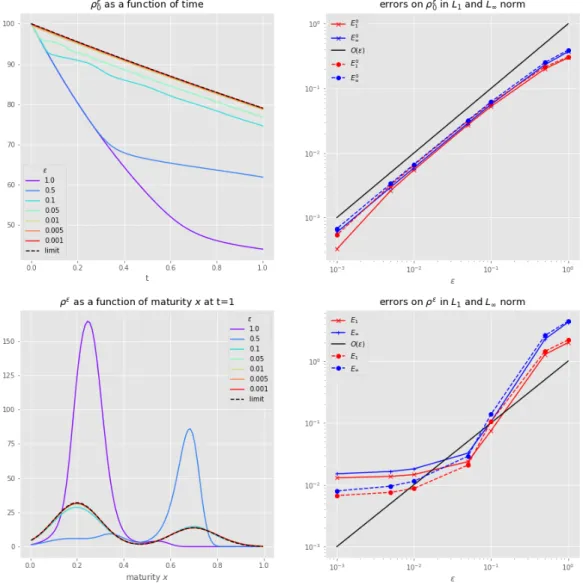


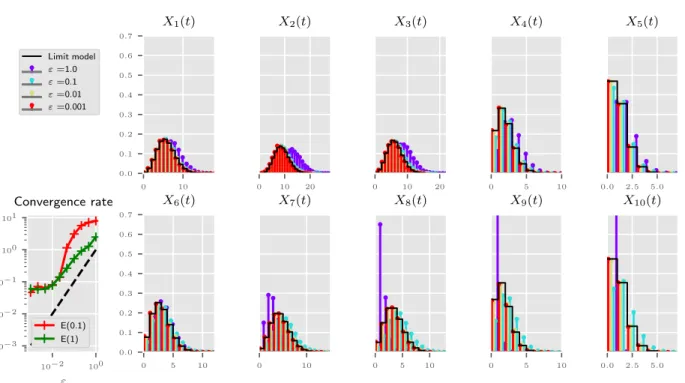
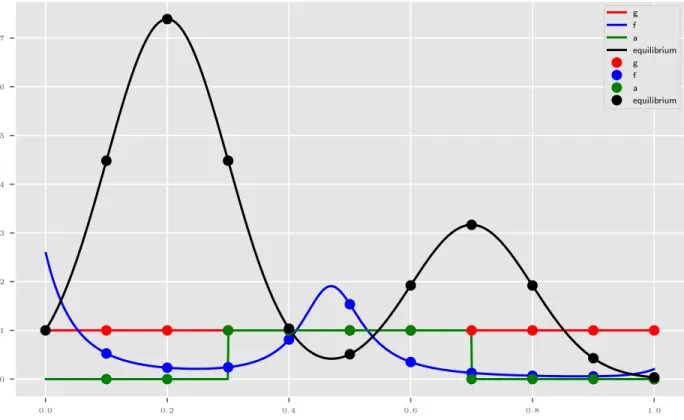
Documents relatifs
Stochastic nonlinear model for somatic cell population dynamics during ovarian follicle activation, (submitted) arXiv :1903.01316.. Stochastic nonlinear model for somatic
Useful in uncertainty quantification (UQ) : provide more reliable ensemble forecasts (EF) and more effiecient spread for ensemble data assimilation (v.s.. perturbations of
0.4.2 Chapitre 2 : Simulations num´ eriques de l’´ equation de Burgers non visqueuse avec des conditions aux limites p´ eriodiques et un terme source stochastique 6 0.4.3 Chapitre 3
This is a coupling of a PDE for the motion of the regular flow, with vorticity ω(t, · ) uni- formly integrable and bounded as in Yudovich’s theorem, and a system of ODEs for the
Les grains de quartz analysés par microscope électronique révèlent deux agents de transport caractérisant ces dépôts ; dans le premier stade (ensemble inférieur et médian),
République Algérienne Démocratique et Populaire Ministère de l'Enseignement Supérieur et de la Recherche Scientifique.. Faculté des Sciences de l ' Ingénieur
Des théories telles que la théorie classique des stratifiés (CLT), la théorie de déformation de cisaillement du premier d'ordre (FSDT) , les théories à ordre élevé (HOTS) et
Using in vitro human trabecular meshwork cells, an in vivo mouse model, and ex vivo human eyes, our study dem- onstrates the potential of clustered regularly interspaced
