HAL Id: inserm-02158503
https://www.hal.inserm.fr/inserm-02158503
Submitted on 18 Jun 2019
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
mammalian auditory sensory epithelium
Ting Zhang, Jinshu Xu, Pascal Maire, Pin-Xian Xu
To cite this version:
Ting Zhang, Jinshu Xu, Pascal Maire, Pin-Xian Xu. Six1 is essential for differentiation and patterning of the mammalian auditory sensory epithelium. PLoS Genetics, Public Library of Science, 2017, 13 (9), pp.e1006967. �10.1371/journal.pgen.1006967�. �inserm-02158503�
Six1 is essential for differentiation and
patterning of the mammalian auditory
sensory epithelium
Ting Zhang1, Jinshu Xu1, Pascal Maire2, Pin-Xian Xu1,3*
1 Department of Genetics and Genomic Sciences, Icahn School of Medicine at Mount Sinai, New York, New York, United States of America, 2 INSERM U1016, Institut Cochin, Paris, France; CNRS UMR 8104, Paris, France; Universite´ Paris Descartes, Sorbonne Paris Cite´, Paris, France, 3 Developmental and Regenerative Biology, Icahn School of Medicine at Mount Sinai, New York, New York, United States of America
*pinxian.xu@mssm.edu
Abstract
The organ of Corti in the cochlea is a two-cell layered epithelium: one cell layer of mechano-sensory hair cells that align into one row of inner and three rows of outer hair cells interdigi-tated with one cell layer of underlying supporting cells along the entire length of the cochlear spiral. These two types of epithelial cells are derived from common precursors in the four- to five-cell layered primordium and acquire functionally important shapes during terminal differ-entiation through the thinning process and convergent extension. Here, we have examined the role of Six1 in the establishment of the auditory sensory epithelium. Our data show that prior to terminal differentiation of the precursor cells, deletion of Six1 leads to formation of only a few hair cells and defective patterning of the sensory epithelium. Previous studies have suggested that downregulation of Sox2 expression in differentiating hair cells must occur after Atoh1 mRNA activation in order to allow Atoh1 protein accumulation due to antagonistic effects between Atoh1 and Sox2. Our analysis indicates that downregulation of Sox2 in the differentiating hair cells depends on Six1 activity. Furthermore, we found that Six1 is required for the maintenance of Fgf8 expression and dynamic distribution of N-cad-herin and E-cadN-cad-herin in the organ of Corti during differentiation. Together, our analyses uncover essential roles of Six1 in hair cell differentiation and formation of the organ of Corti in the mammalian cochlea.
Author summary
Auditory sensory hair cells and surrounding supporting cells are derived from common prosensory progenitors, which undergo rearrangements through intercalation to achieve extension and establish the mosaic structure between hair and supporting cells. Hair cells are susceptible to damage from a variety of insults and are unable to regenerate. Through temporal deletion of Six1 in the developing cochlea, we found that Six1 activity is crucial for proper hair cell fate specification and for the regulation and maintenance of the spatio-temporal pattern of Sox2, Fgf8 and E- and N-cadherins during differentiation. Our data
a1111111111 a1111111111 a1111111111 a1111111111 a1111111111 OPEN ACCESS
Citation: Zhang T, Xu J, Maire P, Xu P-X (2017)
Six1 is essential for differentiation and patterning of the mammalian auditory sensory epithelium. PLoS Genet 13(9): e1006967.https://doi.org/10.1371/ journal.pgen.1006967
Editor: Ping Chen, Emory University School of
Medicine, UNITED STATES
Received: March 10, 2017 Accepted: August 8, 2017 Published: September 11, 2017
Copyright:© 2017 Zhang et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability Statement: All relevant data are
within the paper and its Supporting Information files.
Funding: This research was funded by the NIH
RO1DC014718 and New York STEM contract C029566 to PXX. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared
uncover novel roles of Six1 in hair cell differentiation during the formation of the organ of Corti.
Introduction
In response to a variety of signals, the prosensory progenitors in the floor of the mammalian cochlear duct enter terminal mitosis and then differentiate into a mosaic of mechanosensory hair cells (one row of inner and three rows of outer hair cells) interdigitated with several sub-types of nonsensory supporting cells, including inner border, inner phalangeal, inner and outer pillar and three rows of Deiters’ cells aligned in a medial-to-lateral direction. Failure to correctly produce or maintain these epithelial cells in the organ of Corti causes deafness. Understanding how hair cell morphogenesis is regulated has significant clinical implications, as hair cells are susceptible to damage from a variety of insults and are unable to regenerate.
The cochlea develops from the ventral portion of the otocyst, which elongates and begins to coil at E12 to reach a full 1.5 turns of the cochlear duct by E17.5 [1]. The prosensory progenitor cells proliferate to expand, and after reaching a defined number, exit the cell cycle from apex toward base between E12.5 to E14.5 to form a four- to five-cell layered non-proliferating pre-cursor domain–the primordial organ of Corti, which is marked by expression of p27Kip1[2,3]. Soon after their cell cycle exit, the precursors initiate cell-type specific terminal differentiation near the base toward apex from E14.5 and undergo unidirectional cellular intercalation move-ment called convergent extension to form the two layers of epithelial cells, a lumenal layer of hair cells and a basal layer of supporting cells [3–5]. The Sox family transcription factor Sox2 is known to specify the precursor cells [6]. As distinct cell types undergo their specific differenti-ation in the precursor primordium, Sox2 shows a differential pattern of expression that is highly maintained in supporting cells through adulthood but downregulated in hair cells, which are induced by the basic helix-loop-helix (bHLH) transcription factor Atoh1 [7]. Cur-rent in vitro experimental evidence suggests that Atoh1 and Sox2 may have a mutually antago-nistic relationship, in which Sox2 expression represses Atoh1-induced hair cell formation and expression of Atoh1 in hair cells leads to downregulation of Sox2 [8–11]. However, whether Atoh1 directly antagonizes Sox2 activity in vivo and how Sox2 is downregulated in the differ-entiating hair cells remain unclear. Moreover, despite extensive research on identifying factors that are important for hair cell morphogenesis, how these individual factors interact to gener-ate different types of epithelial cells with distinct shapes and functions in the organ of Corti is still poorly understood. It is even more unclear how these interactions are precisely regulated to induce robust epithelial morphogenesis of the cochlea.
We have recently shown in the cochlear explant that Six1 of the homeodomain protein Six/ So (Sine oculis) family interacts with Eya1 of the phosphatase-transcriptional coactivator Eya (Eyes absent) family to form a key transcriptional complex to activateAtoh1 expression to
induce a hair cell fate by interacting with Sox2 [11].Six1 is expressed in the otic placodal
ecto-derm as early as E8.75 and its expression becomes restricted to the ventral region of the otocyst where inner ear sensory organs form [12,13]. The importance ofSix1 in inner ear development
has been demonstrated by loss-of-function studies in mice and humans [12–15]. Mutations in the humanSIX1 gene cause sensorineural hearing loss [15], and the inner ear defects include either no or an undercoiled cochlea and absence or truncation of vestibular organs [16]. Loss of
Six1 in mice leads to an early arrest of inner ear development at the otocyst stage [12,13]. More recently, a mouse model (Catweasel) carrying a novel point mutation (Cwe) within the Six1
Cwe/Cwe homozygous animals have severely truncated cochlea and semicircular canals [14]. Although the levels of Six1 expression have not been measured inCwe/Cwe animals, the nature
of the inner ear defects associated with this mutation indicates that it is a hypomorphic allele of
Six1. During later stages of inner ear morphogenesis, strong Six1 expression is maintained in
the differentiating hair cells [12,13]. However, despite the absolute necessity ofSix1 for inner
ear development, it remains unknown how Six1 acts to drive sensory hair cell formation and the patterning of the organ of Corti.
In the present study, we used tamoxifen-inducible Cre mice to conditionally deleteSix1
after cochlea duct outgrowth to specifically investigate its potential role in auditory sensory epithelium development. Our data provides novel evidence to support a model in which Six1 serves as a critical factor for hair cell fate induction, differentiation and formation of the audi-tory sensory epithelium.
Results
Six1 conditional mutant mice show defects in proliferation of the
progenitor cells that give rise to the entire organ of Corti in the cochlea
To specifically investigate the role ofSix1 during the specification of sensory epithelial
primor-dium in the developing cochlea, we used an inducible system to temporarily deleteSix1 after
cochlear duct outgrowth by crossing the conditionalSix1floxmice [17] withEya1CreER[18] or
Sox2CreER[19] and administering tamoxifen from E11.5 to E12.5 before the sensory precursor cells exit cell cycle. Lineage tracing usingR26RLacZreporter confirmed that one dose of tamoxi-fen administration at E11.5 inducedEya1CreER-lineage traced cells in the GER and all cells in the organ of Corti at P0 (S1A Fig) [20]. Similarly, previous studies have shown that tamoxifen treatment at E11.5 and E12.5 inducedSox2CreER-lineage traced cells in the GER and all cells in the organ of Corti as well as those in the vestibular organs [21]. Next, we confirmed whether tamoxifen administration at E11.5–12.5 specifically deletes Six1 function in only hair cell pre-cursors or also supporting cell prepre-cursors usingEya1CreERorSox2CreER. Immunostaining with anti-Six1 revealed that Six1 is widely expressed in the cochlear epithelium at E12.5–13.5 (S1B Fig), butSix1 CKO (Eya1CreER;Six1fl/florSox2CreER;Six1fl/fl) showed a significant reduction in the levels of Six1 within the sensory region 1–2 days after tamoxifen treatment (S1B Fig). At E17.5, Six1 antibody appeared to label not only hair cells but also supporting cells and the flanking GER/LER cells (S1C Fig), Six1 expression was lost in the hair cells inSix1 CKO
cochlea using eitherEya1CreERorSox2CreERas a deletor (S1D and S1E Fig). In addition, Six1 expression levels in some supporting cells adjacent to the GER also appeared to be reduced (S1D and S1E Fig). However, Six1 signal was strongly maintained in GER cells, some support-ing cells and LER cells in the mutant, suggestsupport-ing that the expression of Six1 in those precursor cells is activated before removal of Six1 and maintained even after tamoxifen induction. Based on these data, we conclude that bothEya1CreERandSox2CreERare able to specifically delete Six1 function in the hair cell precursors within the organ of Corti.
To assess the effect of loss ofSix1 function between E11.5–12.5 on the establishment of the
prosensory epithelial domain in the cochlea, we harvested inner ears from embryos at E14.5 and found thatSix1Cko/Cko(Eya1CreER;Six1fl/florSox2CreER;Six1fl/fl) inner ears were noticeably smaller in size compared to wild-type,Eya1CreERorSox2CreERlittermate controls (n = 6
embryos for each genotype;S2A Fig). We performed co-immunostaining of E14.5 cochleae with anti-Sox2 antibody to label all prosensory progenitors and p27Kip1to mark postmitotic precursors in the nascent organ of Corti (Fig 1). In wild-type embryos, the cochlea had already reached more than one turn and most Sox2+progenitors had exited the cell cycle to become p27Kip1+along the entire cochlear duct (Fig 1A). InEya1CreERorSox2CreERlittermates, the
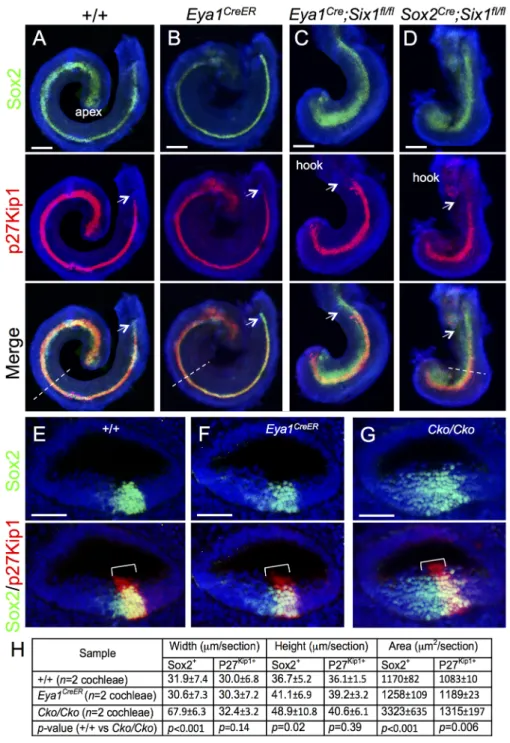
Fig 1. Deletion of Six1 in the developing cochlea using Eya1CreERor Sox2CreERleads to shortened and
thickened prosensory primordium. Cochleae were dissected from E14.5 embryos given tamoxifen at E11.5 (9 am) and E12.5 (9 am) and processed for whole-mount (A-D) or section (E-G) immunostaining with anti-Sox2 (green) and -p27Kip1(red). Hoechst was used for nuclear-counter staining. (E-G) Section collected from mid-cochlear duct in wild-type, Eya1CreERor Sox2CreER;Six1fl/fl(Cko/Cko) littermates as indicated by
dashed line in A, B, D respectively. Bracket indicates p27Kip1-positive prosensory domain within the cochlea
epithelium and its width on mediolateral axis is comparable between control and mutant littermates. (H) Spatial calibration of Sox2+and p27Kip1+width, height and square area and value represents average number (±standard deviations) per section (6μm) (seeMethodsfor calibration). P-value was measured for +/+ and Cko/Cko using Two-tailed Student’s t-test. Scale bars: 200μm in A-C and 40μm in D,E.
cochlea development appeared indistinguishable from that in wild-type controls at this stage (n = 6 embryos,Fig 1B). However, inSix1 CKO littermates, shortening of the cochlear duct
was evident at E14.5 (Fig 1C and 1D), and its length was comparable to that of the E12.5 con-trol embryos [1]. While the Sox2+domain expanded medially inSix1 CKO samples along the
length of the cochlear duct, most of Sox2+cells still underwent cell-cycle exit to form the p27Kip1+non-proliferating domain, which almost reached the base (Fig 1C). However, some
Six1 CKO embryos had fewer p27Kip1+cells in the basal end (n = 5 out of 11 embryos, arrows
inFig 1D, compare to 1A-C). This suggests that not all prosensory progenitors in the base completed their cell-cycle exit to become non-proliferating precursors in the CKO. Immunos-taining for Sox2/p27Kip1on sections showed medially widened Sox2+domain in the CKO (Fig 1E–1G). Statistical analysis of the width, height and area of Sox2+and p27Kip1+domain (n = 2
cochleae and 15 sections per cochleae) confirmed that the Sox2+domain within the CKO cochlear epithelium is widened compared to that in littermate controls (Fig 1H).
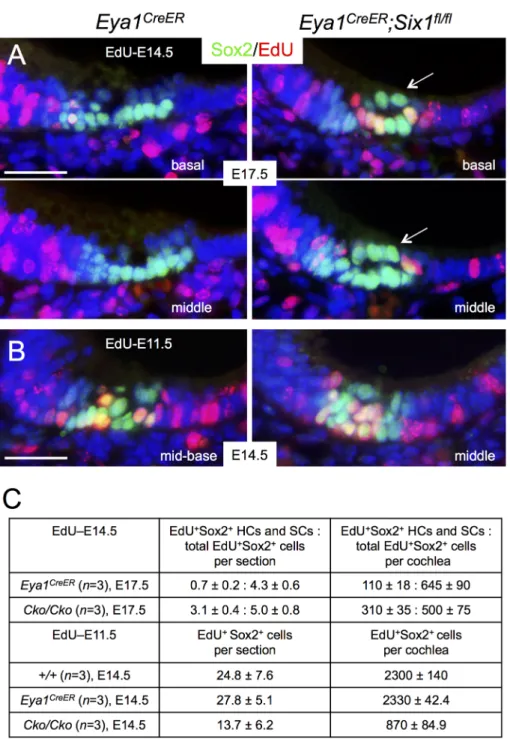
To confirm that p27Kip1-negative Sox2+progenitors in the base of the CKO cochlea are indeed proliferative progenitors, we injected the mitotic tracer 5-ethynyl-2’-deoxyurindine (EdU) at E14.5 and harvested inner ears at E17.5. Co-immunostaining for EdU and Sox2 and quantitative cell counting confirmed that there were more EdU-incorporated hair cells and supporting cells, majority of which was located in the base, in the CKO mutant than in control littermates (Fig 2A and 2C). More EdU-incorporated p27Kip1+cells within the sensory epithe-lium were also observed in the base ofSix1 CKO cochlea (S2C and S2D Fig), while theSix1
CKO inner ears were also smaller in size compared to littermate controls (S2B Fig). Together, these data suggest that there is a slight delay in the sensory epithelium development in theSix1
CKO.
Next, we asked whether defective cochlear elongation between E12.5 and E14.5 prior to ter-minal differentiation of the precursor cells inSix1 CKO mutant might be, at least in part, due
to defective cell proliferation by co-injecting EdU together with tamoxifen at E11.5. Immunos-taining and quantitative analysis indicated that EdU-incorporated Sox2+cells were reduced to ~37% of those in the littermate controls at E14.5 (Fig 2B and 2C). TUNEL assay revealed that the number of apoptotic cells was only mildly increased in the CKO at E12.5–14.5 compared to the number in littermate controls (S3 Fig). Together, these data indicate that Six1 activity is necessary for normal cell proliferation of the epithelial progenitors and cochlear growth.
Loss of Six1 alters the spatiotemporal pattern of Sox2 expression during
differentiation of the organ of Corti
Sox2 specifies prosensory progenitors and is expressed in all progenitor cells at the early stages, but later during differentiation its expression is downregulated in hair cells and becomes restricted to the supporting cells in the organ of Corti [6,8]. By E17.5, Sox2 levels in hair cells located in the basal and medial cochlear regions are normally downregulated in comparison to its high expression levels in the supporting cells (Fig 2A). However, we noticed that high levels of Sox2 are maintained in all cells within theSix1 CKO organ of Corti (arrows,Fig 2A). This led us to speculate that Six1 may regulate the spatiotemporal pattern of Sox2 expression in the organ of Corti during differentiation. To rule out the possibility that the high levels of Sox2 in the cells within the lumenal layer in the CKO organ of Corti is due to developmental delay and confirm that those high Sox2+cells are indeed hair cells, we harvested cochlea 1–2 days later at E18.5-P0 after tamoxifen administration at E11.5–12.5 and performed immunostaining for Sox2 and Myo7a, a marker specific for differentiating hair cells. While strong Sox2 expression is maintained in supporting cells through adulthood, relatively low Sox2 activity is detectable in GER (greater epithelial ridge) cells flanking the inner hair cells at E18.5 (Fig 3A and 3B). In
Six1 CKO, the cochlear duct not only appeared wider and thicker with discontinuation of
Sox2+domain in the base compared to that in wild-type controls (arrows,Fig 3A;n = 6), but
also was shortened to 0.75- to 1-turn. Along with cochlear elongation between E14.5 to E18.5, Fig 2. Altered cell proliferation in Six1 CKO (Eya1CreER) cochlea epithelium. (A) Immunostaining for Sox2 (green) and EdU (red) on sections of basal and middle regions of cochleae from E17.5 embryos given EdU at E14.5 and tamoxifen at E11.5–12.5. Arrows point to high levels of Sox2 in cells within the lumenal layer. (B) Immunostaining for Sox2 (green) and EdU (red) on sections of middle region of cochleae from E14.5 embryos given tamoxifen at E11.5–12.5 and EdU at E11.5. (C) Number of EdU+Sox2+cells per section (6μm) or EdU+cells per cochlea (seeMethodsfor quantification). P-value was measured for +/+ and Cko/Cko using
Two-tailed Student’s t-test: p<0.001 for E17.5 (EdU at E14.5) and p = 0.003 for E14.5 (EdU at E11.5). Scale bars: 30μm in A,B.
hair cell differentiation occurs near the mid-base and reaches the basal end and apex in a medial-to-lateral gradient to form one row of inner and three rows of outer hair cells along the entire length of the cochlea by E18.5 as marked by Myo7a (Fig 3B). The hair cells are interdigi-tated by distinct subtypes of specialized supporting cells: one row of inner border cells and one row of inner phalangeal cells surrounding the inner hair cells, two rows of pillar cells (one row of inner and one row of outer pillar cells) lining the space between the inner and outer hair cells–the tunnel of Corti–and three rows of Deiters’ cells associated with the outer hair cells. In
Six1 CKO cochlea, Myo7a+hair cells were indeed present but they appeared irregularly with only one cell toward the base and more than four cells toward the apex. However, high levels of Sox2 expression were still maintained in Myo7a+hair cells, as in the supporting cells that also appeared to be aligned irregularly. Furthermore, lower Sox2 activity appeared to expand medially into GER cells in the mutant (arrows,Fig 3B).
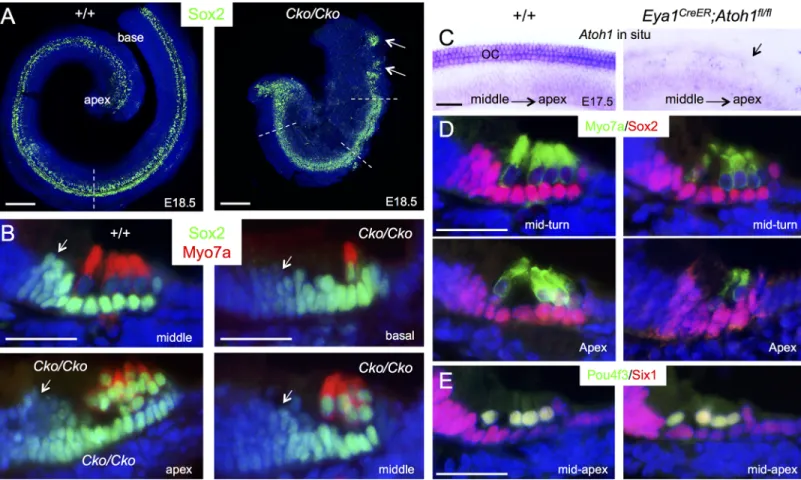
The observation of high levels of Sox2 expression inSix1 CKO hair cells was a surprising
finding because it has been argued that Atoh1 is involved in the downregulation of Sox2. To test this further, we examined the dependence of Sox2 levels on Atoh1 in hair cells by deleting
Atoh1 from E14.5–15.5 using Eya1CreER. In situ hybridization of cochlea at E17.5 confirmed Fig 3. Downregulation of Sox2 in differentiating hair cells is disrupted in Six1 CKO. (A) Immunostaining for Sox2 (green) on whole-mount and (B) Sox2 (green)/Myo7a (red) on sections of cochleae from wild-type or Six1 CKO (Eya1CreER;Six1fl/fl) littermate embryos at E18.5 (given tamoxifen at E11.5–12.5).
Cochlear section region was indicated by dashed line in A. Arrows point to Sox2 expression in the GER cells flanking the inner hair cells. (C) Atoh1 in situ hybridization showing Atoh1 expression in the organ of Corti in wild-type embryos at E17.5 and absence of Atoh1 in Atoh1fl/fl;Eya1CreERlittermate embryos (arrow) given tamoxifen at E14.75–15.5. (D) Immunostaining for Myo7a (green) and Sox2 (red) on cochlear sections from E17.5 wild-type or Eya1CreER; Atoh1fl/fllittermate embryos given tamoxifen at E14.75–15.5. (E) Immunostaining for Pou4f3 (green) and Six1 (red) on cochlear sections from E17.5 wild-type
and Eya1CreER;Atoh1fl/fllittermate embryos given tamoxifen at E14.75–15.5. Scale bars: 200μm in A; 30μm in B,D,E; 40μm in C.
deletion ofAtoh1 in the differentiating hair cells (Fig 3C). However, immunostaining for Sox2, Six1 and Pou4f3 revealed no detectable changes inAtoh1 CKO (Fig 3D and 3E), which was consistent with previous observations detected by western blot and in situ hybridization for these genes [22]. Thus, deletion ofAtoh1 in differentiating hair cells does not lead to
upregula-tion of Sox2. Based on these data, we conclude that Six1 activity is crucial for downregulaupregula-tion of Sox2 in the differentiating hair cells during cochlear development.
Six1 is required for hair cell fate specification in the cochlea
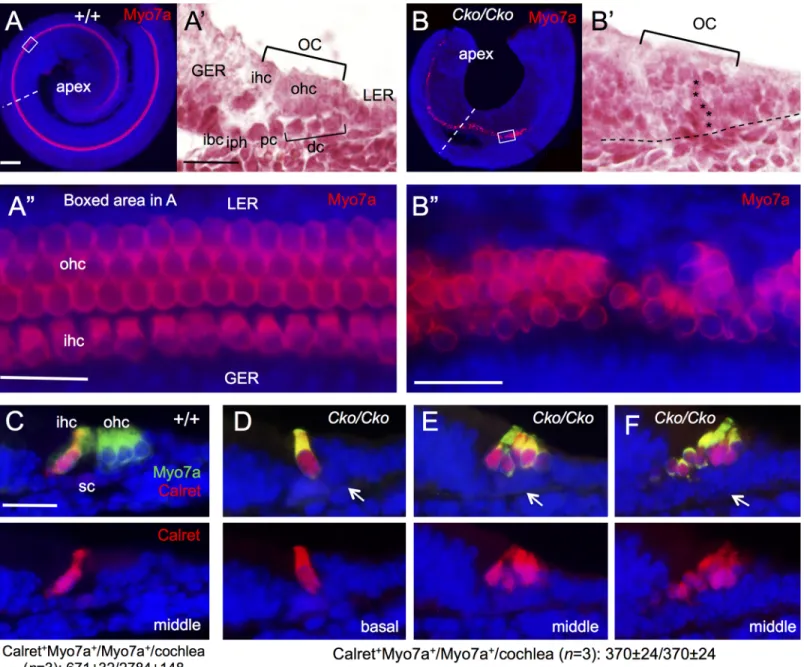
As inner hair cells differentiate prior to outer hair cells, we next sought to characterize whether the Myo7a+hair cells observed inSix1 CKO cochlea are inner hair cells, outer hair cells or
both. Whole-mount immunostaining of cochlea at E18.5 revealed one row of inner and three rows of outer hair cells along the entire length of the cochlea in wild-type control (Fig 4A and 4A”). In theSix1 CKO, while the length of cochlea was shortened to ~0.75- to 1-turn, Myo7a+
cells extended to the apical end but were missing in the base (n = 6;Fig 4B). This suggests that hair cell differentiation toward the basal end fails to occur.
Higher magnification analysis showed that hair cells that had formed inSix1 CKO had
abnormal morphology and irregular alignment with an uneven numbers of hair cells on the mediolateral axis, ranging from one to multiple cells (Fig 4B”). As seen on sections, the organ of Corti consists of two layers of epithelial cells, a lumenal layer of hair cells and a basal layer of supporting cells (Fig 4A’) flanked by nonsensory epithelial cells in the GER/LER (greater/lesser epithelial ridge). In contrast,Six1 CKO organ of Corti is retained as a four- to five-cell layered
epithelium (Fig 4B’), which is almost comparable to the non-proliferating precursor domain in E14.5 control embryos. This clearly indicates a defect during terminal differentiation of the p27Kip1+precursor cells in the mutant. Analysis of vestibular sensory organs showed largely reduced utricular and saccular macula with fewer hair cells and no hair cells in crista ampul-laris in all three semicircular canals (S4 Fig).
Interestingly, all Myo7a+cells inSix1 CKO cochlea are positive for Calretinin (Fig 4D–4F), a marker specific for inner hair cells (Fig 4C), suggesting that the hair cells developed in the CKO cochlea treated with tamoxifen between E11.5–12.5 might be inner hair cells. Quantita-tive counting revealed that the total number of hair cells inSix1 CKO cochlea (Myo7a+ Calreti-nin+cells/total Myo7a+cells = 370±24/370±24 per cochlea; n = 3) decreased to ~53% of the total number of inner hair cells in wild-type cochlea (Calretinin+Myo7a+cells/total Myo7a+ cells = 671±32/2784±148 per cochlea; n = 3). Thus, the absence of outer hair cell formation in
Six1 CKO is not likely due to a conversion of outer hair cells to inner hair cells, but rather
caused by a failure of activation of the outer hair cell differentiation program.
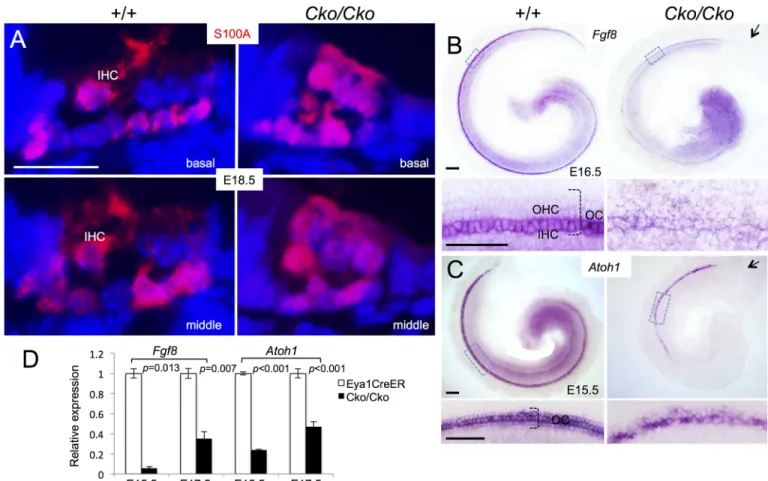
To further confirm our observation, we immunostained for the calcium-binding protein S100A, which labels inner hair cells, inner phalangeal cells and Deiters’ cells [23,24] (Fig 5A). InSix1 CKO cochlea, S100A labeled not only all hair cells but also all supporting cells. This
fur-ther suggests that the remaining hair cells in the CKO might be inner hair cells.
In situ hybridization withFgf8 riboprobe, a marker specific for inner hair cells [25], re-vealed strongFgf8 expression in the inner hair cells in E16.5 cochlea (Fig 5B). However,Fgf8
expression was decreased inSix1 CKO littermates and appeared in the remaining hair cells at
E16.5 (Fig 5B). Quantitative real-time RT-PCR (qRT-PCR) confirmed a large reduction of
Fgf8 expression in the mutant at E15.5 and E17.5 (Fig 5D). Together, these results suggest that Six1 may regulate the maintenance ofFgf8 expression in the inner hair cells.
In situ hybridization confirmed thatAtoh1 mRNA is expressed in the hair cells of Six1
CKO cochlea at E15.5, at whichAtoh1 expression has not yet reached its apex in both controls
CKO than in the control littermates (Fig 5C), and this reduction was confirmed by qRT-PCR (Fig 5D). Thus, whileAtoh1 expression is induced in the postmitotic precursors, which only
differentiate into inner hair cells inSix1 CKO treated with tamoxifen between E11.5–12.5, Six1
may play a role in controlling the maintenance or upregulation ofAtoh1 in hair cells during
differentiation.
As outer hair cells differentiate more than one day after the onset of inner hair cell differen-tiation, we next sought to further clarify our observation by administering tamoxifen more than one day later between E12.75-E13.5 or E13.5–14.5 to examine whether outer hair cells Fig 4. Lack of outer hair cell formation in Six1 CKO (Eya1CreER) cochlea. (A,B) Immunostaining for Myo7a of cochlea from E18.5 embryos given
tamoxifen at E11.5–12.5. (A’,B’) H&E staining of section from middle region of cochlea indicated by dashed line in A or B respectively. (A”,B”) Higher magnification of boxed region in A or B respectively. (C-F) Co-immunostaining on sections for Myo7a (green) and Calretinin (red). Abb.: dc, Deiters’ cells; GER, greater epithelial ridge; ibc, inner border cell; ihc, inner hair cells; iph, inner phalangeal cell; oc, organ of Corti; ohc, outer hair cells; pc, pillar cells; sc, supporting cells. Scale bars: 200μm in A and B; 50μm in A’,B’ and C-F; 30μm in A” and B”.
also form in the mutant. Analysis of E18.5 cochleae treated with tamoxifen between E12.75– 13.5 showed that theSix1 CKO cochlea reached a full 1.5 turns, and that four rows of hair cells
formed in the basal turn, but revealed a pattern of severity that parallels the normal process of hair cell differentiation, with outer rows more affected than inner rows in the medial turn, and no outer hair cells in the apex (n = 6,Fig 6A). A similar observation was obtained in P0 cochleae treated with tamoxifen between E13.5–14.5 (n = 6,Fig 6B). All hair cells formed in
Six1 CKO cochlea also displayed abnormal morphology and maintained high levels of Sox2
(Fig 6B). These data provide additional evidence that Six1 activity is necessary for hair cell fate specification and downregulation of Sox2 in the differentiating hair cells.
Alteration in cell morphology and distribution of cadherins in Six1 CKO
organ of Corti
We failed to observe S100A-negative pillar cells on all sections from theSix1 CKO cochleae
(n = 3,Fig 5A), suggesting that the pillar cells are not formed in the mutant. We therefore fur-ther investigated whefur-ther loss ofSix1 also results in loss of supporting cell subtypes, using
Fig 5. Reduced Fgf8 expression in Six1Cko/Ckomutant (Eya1CreER;Six1fl/flgiven tamoxifen from E11.5–12.5). (A) Immunostaining for S100A (red) on sections of cochleae from E18.5 wild-type or Six1Cko/Ckolittermate embryos. (B) Whole-mount in situ hybridization with Fgf8 ribo-probe showing Fgf8 expression in inner hair cells (IHC) in wild-type control littermates and decreased expression in remaining hair cells in Six1Cko/Cko
littermates at E16.5. Lower panels are higher magnification of boxed areas. (C) Whole-mount in situ hybridization of E15.5 cochleae showing Atoh1 mRNA expression in the hair cells in Six1Cko/Ckocochlea. Arrow in B,C points to lack of Fgf8 or Atoh1 expression in the basal end of cochlea in Six1Cko/Cko. Lower panels are higher magnification of boxed areas. (D) Fgf8 and Atoh1 mRNA expression were examined by qRT-PCR in Eya1CreER and Six1 CKO littermates at E15.5 and E17.5 (given tamoxifen at E11.5–12.5). Two-tailed Student’s t-test was used for statistical analysis. Scale bars: 20μm in A,B, 100μm in upper panels of C,D and 50μm in lower panels of C,D.
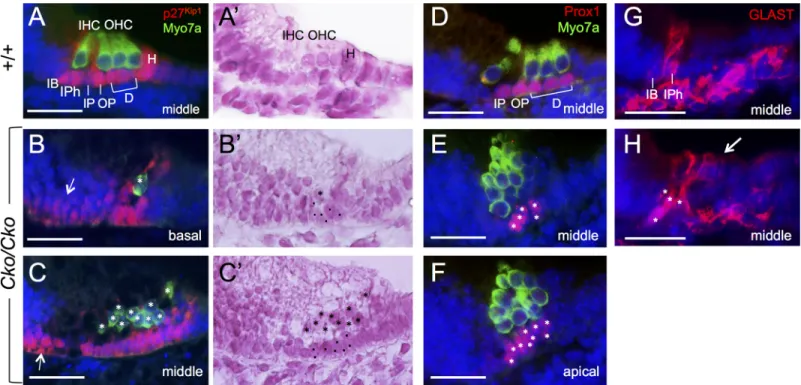
specific marker gene analysis. Despite irregular shape, all Sox2+supporting cells underlying the hair cells inSix1 CKO cochlea were positive for p27Kip1at E18.5 (n = 5,Fig 7B and 7C), whose expression is normally restricted to all supporting cells, including Hensen’s cells flank-ing the outermost outer hair cells (Fig 7A). Similar to Sox2, p27Kip1expression also showed medial expansion to the flanking GER cells (arrows,Fig 7B and 7C,n = 3). However, those
cells expressed neither hair cell markers nor other supporting cell markers. For example, Prox1, which is expressed in pillar cells and Deiters’ cells (Fig 7D) and thought to act down-stream of Sox2 [8], was found to be expressed in supporting cells underlying the hair cells but not in adjacent GER cells inSix1 CKO cochlea at E18.5 (n = 4,Fig 7E and 7F). However, the Prox1+cells in the CKO failed to align into a characteristic one-cell layer, and there were five or more in the basal to middle region (Fig 7E) and up to ten Prox1+cells underlying the two-to three-cell layered hair cells two-toward the apex (Fig 7F). Inner border and inner phalangeal cells labeled by Glutamate-aspartate transporter (GLAST) (Fig 7G) were detectable in the medial region of the organ of Corti inSix1 CKO (n = 4). However, in contrast to their apical
process that only surrounds the inner hair cells in the wild-type littermates (Fig 7G), these GLAST+cells in the mutant appeared to make apical process that surrounded all hair cells (arrow,Fig 7H).
As Prox1 is expressed in both inner and outer pillar cells, we next used inner pillar cell spe-cific marker p75NTRto further clarify the presence of inner pillar cells in the CKO. Indeed, inner pillar cells labeled by p75NTRwere present but showed changes in cell shape and cell Fig 6. Temporal deletion of Six1 between E12.75–13.5 blocks hair cell induction. (A) Whole-mount Myo7a staining of E18.5 cochleae showing four rows of hair cells along the entire cochlea in Eya1CreERmice but extra inner hair cells present from middle toward apex (arrows). In Six1 CKO
cochlea, the organ of Corti appears narrower and hair cells show abnormal morphology with gradually decreased outer rows of hair cells from four in the base and one row in the apex. (B) Co-immunostaining for Myo7a and Sox2 on sections from P0 cochleae treated with tamoxifen between E13.5– 14.5 showing one inner and three outer hair cells in the base, one inner and two outer hair cells in the middle and only one hair cells in the apex in Six1 CKO. All Myo7a+hair cells are also Sox2+in the CKO mutant. Scale bars: 30μm.
contacts on the lumenal surface. During cochlear elongation, rearranging epithelial cells shrink junctions that are oriented perpendicular to the axis of extension and subsequently resolve such shrinkage to restore more isodiametric shapes. In control animals at E17.5, inner pillar cells in the basal region are aligned in a single row with stable cellular contacts (long junction) (Fig 8A). Dynamic remodeling of cellular contacts was seen toward the less differentiated api-cal region (arrow inFig 8A). InSix1 CKO, the inner pillar cells showed changes in morphology
and there were three or more inner pillar cells in contact (arrows,Fig 8B,n = 5) and cellular
contact shrinkage (arrowhead,Fig 8B) throughout the entire cochlea, indicating a clear defect in cellular rearrangement in the absence ofSix1.
Further examination also uncovered morphological alteration in hair cells inSix1 CKO.
During differentiation, hair cells form actin-rich V-shaped stereociliary bundle with graded heights that are all individually aligned and point in the same direction toward the lateral side of the organ of Corti. This polarization process is initiated by the migration of the centrally positioned kinocilium to the periphery from ~E16.5 [26]. At P0, the uniform orientation of hair cells and their interdigitation with nonsensory supporting cells on the lumenal surface with asymmetric and lateral distribution of kinocilium as outlined by F-actin and acetylated tubulin staining respectively were evident throughout the entire organ of Corti in wild-type controls (n = 6,Fig 8C). However, in absence ofSix1 function, we found that individual hair
cell orientation was severely affected (n = 6,Fig 8C). Collectively, our results demonstrate that absence ofSix1 not only limits the normal extension of the cochlear duct, but also results in
significant defects in cell shape within the plane of cochlear sensory epithelial sheet during ter-minal differentiation.
Fig 7. Differentiation and misalignment of supporting cell subtypes in the absence of Six1. Antibody labeling for Myo7a for hair cells and p27Kip1
(A-C), Prox1 (D-F) and GLAST (G,H) for supporting cells in the organ of Corti in wild-type and Six1 CKO (Eya1CreER;Six1fl/fl) littermates given tamoxifen from E11.5–12.5. A’,B’,C’ are A.B,C counter-stained with hematoxylin respectively. Arrow in B,C points to expansion of p27Kip1expression in the GER cells. Asterisks in B,B’,C,C’ point to remaining hair cells and dots point to supporting cells. Asterisks in E,F,H point to supporting cells. Abb.: D, Deiters’ cells; H, Hensen’s cells; IB, inner border cell; IHC and OHC, inner and outer hair cells; IPh, inner phalangeal cells; IP and OP, inner and outer pillar cells. Scale bars: 30μm.
As selective cell adhesion, mediated by cadherins, plays a pivotal role in regulating the shape and topology of the cells in tissue morphogenesis [27], we therefore tested whether loss ofSix1 leads to changes in adhesion by comparing the distributions of cadherins in controls
andSix1 CKO littermates. At E15.5, the organ of Corti has differentiated in the base and
mid-dle, and consists of one row of inner and three rows of outer hair cells interdigitated with non-sensory supporting cells as outlined by phalloidin staining (Fig 8D). N-cadherin is expressed in the cochlea from E14.0 [28] and its distribution is restricted to cells medial to the outer hair cell from E15.5 (Fig 8D). InSix1 CKO, the general integrity of the organ of Corti is not
main-tained and as expected, due to the expanded (more than one-cell) layer of hair cells (Fig 4B’ and 4B”), the organization between hair cells and their interdigitated nonsensory supporting cells is apparently disrupted, similar to that observed at P0 (Fig 8C). InSix1 CKO, N-cadherin
expression was observed in the medial region, but its expression expanded to the lateral region of the organ of Corti (n = 5,Fig 8D). In contrast, E-cadherin is normally restricted to the region of outer hair cells and the region lateral to it from E16.0 (Fig 8E), and its onset of mem-brane localization coincides with the stabilization of cell junctions in the region lateral to the inner pillar cells. However, the levels of E-cadherin at the cell membrane were reduced inSix1
CKO cochlea at E16.0 (n = 4,Fig 8E). By E18.5, relatively lower levels of E-cadherin at the cell membrane were widely detectable in all cells within the sensory epithelium as well as in the GER cells in the CKO (n = 4,Fig 8F). These results indicate that loss ofSix1 function alters the
patterns of N- and E-cadherin distribution in the cochlea and the structure of the organ of Corti.
Discussion
The role ofSix1 in inner ear development has been previously investigated [12–14]. However, due to lack of inner ear formation beyond the otocyst stage inSix1-null mice, the molecular
modules carried out bySix1 in the auditory sensory epithelium remain unknown. In cochlear
explant, we have shown that Six1 forms a complex with Eya1 and Sox2 to activateAtoh1
expression to induce hair cell fate in the GER cells [11]. Here, we for the first time investigated the in vivo requirement ofSix1 in auditory sensory epithelium specification and
differentia-tion. Our analyses indicate that Six1 is crucial not only for proper fate specification but also for proper patterning of the precursor cells in the auditory sensory epithelium, which are neces-sary steps for the formation of the organ of Corti in the cochlea.
Previous studies have shown that, in the otocyst, Six1 promotes both proliferation and sur-vival of the otic epithelial cells [12,13]. However, Six1 activity does not appear to be crucial to cell survival in the developing cochlea as no significant difference in apoptosis was observed in
Six1 CKO. Reduced proliferation in Six1 CKO cochlear epithelium suggests that Six1 plays a
critical role in maintaining the prosensory progenitor cells at proliferative state in order to expand to a certain number, which explains shortened cochlea and a reduced number of hair Fig 8. Alteration in cellular morphology and distribution of N- and E-cadherin in Six1 CKO organ of Corti (tamoxifen at E11.5–12.5, Eya1CreER). (A,B) Inner pillar cells labeled by p75NTRin wild-type cochlea and Six1 CKO littermate cochleae at E17.5. Arrow in A points to dynamic remodeling of cellular contacts toward the less differentiated apical region. Arrows and arrowhead in B point to three or more inner pillar cells in contact and cellular contact shrinkage respectively in Six1 CKO organ of Corti. (C) F-actin (phalloidin) and acetylated tubulin staining showing alteration in cell orientation and alignment in Six1 CKO at P0. (D-F) Lumenal surface views of the cochleae from wild-type, Eya1CreER, and Six1 CKO (tamoxifen at E11.5–12.5, Eya1CreER) (D,E) embryos at E15.5
(D), E16.0 (E) or E18.5 (F). Samples were stained for phalloidin and N-cadherin (D) or E-cadherin (E,F). Arrowheads mark the separation between inner hair cells and outer hair cells. The developing organ of Corti is identified by cellular morphology and cortical actin enrichment in the nascent hair cells. Scale bars: 30μm A,B, 20μm C, and 30μm E-F.
cells observed inSix1 CKO. How does Six1 act to regulate cell proliferation? We found that
Six1 forms a complex with Eya1, N-Myc and c-Myc proteins in E12.5–13.5 cochlea [11,29]. Myc proteins are known to be important for cell proliferation and growth, making it plausible to speculate that Six1 works together with its cofactors such as Eya1 and Myc proteins to regu-late cell proliferation and growth. Defects in cell division and growth before terminal mitosis are likely to lead to shortened cochlea occurring in theSix1 CKO mutant.
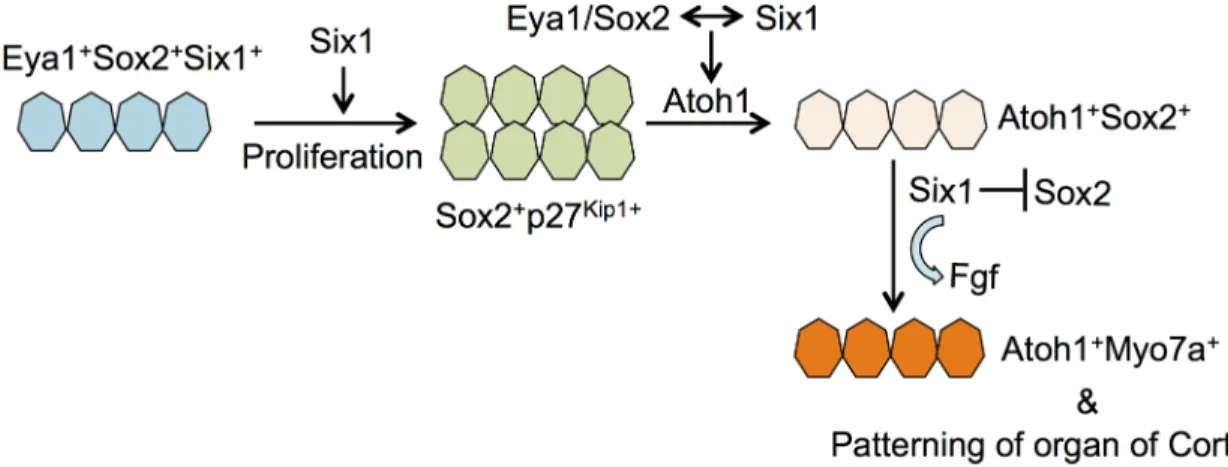
Our observation of the absence of outer hair cell formation in theSix1 CKO mutant
pro-vides the first in vivo evidence supporting a model in which Six1 serves as a key factor for hair cell fate specification (Fig 9). In cochlear explants, we found that Six1 forms a complex with Eya1 and Sox2 to synergistically activateAtoh1 to induce hair cell fate [11]. Given that Eya1 is a transcriptional coactivator interacting with DNA-binding proteins Six1 and Sox2 to transac-tivateAtoh1 expression, and the latter (Sox2) is necessary for Atoh1 activation in vivo [6,11,
30], we have previously proposed a model in which Eya1 bridges Sox2 and Six1 to undergo protein-interaction-dependent and binding sequence-dependent conformational changes to form a compact and active complex capable of transcriptional activation ofAtoh1 [11]. Based on this model, if all three genes are necessary forAtoh1 activation in vivo, deletion of any one
of the three genes would lead to failure to induceAtoh1 expression and hair cell fate
specifica-tion. Deletion ofEya1 in the differentiating hair cells using Atoh1-CreER at E13.5–14.0 fails to
activateAtoh1 and results in the absence of both inner and outer hair cell differentiation in the
apex [11], demonstrating the necessity of Eya1 activity forAtoh1 activation in vivo, which is
induced from ~E13.0 and becomes detectable at ~E13.5 [4,31]. So how do we explain the pres-ence ofAtoh1 expression in the Six1 CKO mutant cochlea? There are two probable
explana-tions. First, as it takes at least ~6 hours for tamoxifen to induce CreER in the nucleus, it is possible that weakAtoh1 expression might have already been induced in some precursors
before complete removal ofSix1 by tamoxifen administration from E11.5–12.5. Such weak Atoh1 expression might have been sufficient to induce hair cell differentiation, which would
also explain why outer hair cells failed to form and why the number of inner hair cells in the
Six1 CKO decreased to ~53% of that of the wild-type control. Second, there may be functional
redundancy with other members of the Six gene family. WhileSix4 mice are normal [32], Six4 is coexpressed with Six1 in otic progenitors in the otocyst [13] and in the cranial placodes, and Fig 9. Model illustrating the role of Six1 in regulating auditory sensory cell development. This study demon-strates that Six1 regulates proliferation of sensory progenitors in the cochlea epithelium. While a direct interaction between Eya1/Six1/Sox2 proteins coordinately regulates Atoh1 expression in cochlear explant, this study provides in vivo evidence supporting a role for Six1 in hair cell fate specification and Atoh1 expression. In differentiating hair cells, our data show that Six1 activity is necessary for downregulation of Sox2 and maintenance of Fgf8 expression.
is known to play redundant roles with Six1 in cranial placode development as well as in several other organ systems [33–39]. Thus, it is possible that Six4 activity in the otic epithelial precur-sors may participate in complex formation with Eya1 and Sox2 to activateAtoh1 in the
prosen-sory precursors. In support of this idea, we found that coexpression of Eya1 and Six4 in cochlear explants can also induce hair cell fate as efficiently as the combination of Eya1 and Six1 [11]. Besides Six4, weak Six2 expression is detected in the cochlear epithelium [40], where Six5 activity may also exist as mutations in human SIX5 [41], SIX1 [15] or EYA1 [42,43] cause Branchio-Oto-Renal or Branchio-Oto syndrome. Thus, Six4, Six2 or Six5 in the prosensory progenitor cells may participate in complex formation with Eya1 and Sox2 for initial Atoh1 activation, but this is clearly insufficient to replace Six1 in upregulating Atoh1 in the differenti-ating hair cells either due to the absence of their expression or low activity in the differentidifferenti-ating hair cells. While the relative ratio of Sox2, Eya1, Six1 and Atoh1 in the precursor cells may be important in specifying hair cells, our data clearly demonstrate that Six1 activity is necessary for specifying hair cell fate in vivo.
The pattern of the outer hair cell defect induced by tamoxifen at later stages between ~E13.0– 14.5 provides additional support for Six1 as a key factor in specifying the gradient pattern of hair cell differentiation longitudinally and laterally in the cochlea. In the normal organ of Corti, hair cell differentiation begins in the mid-base and extends not only laterally, but also towards both the basal end and the apex of the cochlea [4]. The longitudinal pattern of hair cell differentiation is also disrupted in theSix1 CKO, which had no hair cell formation in the basal end (Fig 4B). Thus, Six1 is likely to play a key role in establishing the hair cell developmental program within the auditory sensory epithelial sheet. While detailed characterization of the Six1-regulatory network controlling hair cell development is an ongoing project in the laboratory, our observation of downregulation ofFgf8 in the Six1 CKO suggests that Six1 may regulate Fgf8 signaling pathway
during differentiation. A key piece of supporting evidence for this has been recently obtained from our ChIP-seq analysis for whole-genome mapping of Six1 in the cochlea, which identified a Six1 peak near theFgf8 gene. Characterization of this Six1-bound peak sequence in transgenic
mice by cloning upstream of theHsp68 minimal promoter driven-LacZ reporter cassette flanked
by the H19 insulators has indicated that this enhancer fragment is a cochlear hair-cell specific enhancer (J. Li, E. Loh, J. Xu, T. Zhang, L. Shen and P-X. Xu, manuscript in preparation). While detailed transgenic and mutational analyses of the Six1-binding site within this enhancer ofFgf8
in vivo are still currently underway, our data suggest thatFgf8 is a direct in vivo target of Six1 in
the cochlea.
In the organ of Corti, Fgf8 is expressed in the inner hair cells and previous studies have sug-gested that Fgf8 may act through its receptor Fgfr3 to regulate pillar cell development [25,44,
45]. InSix1 CKO, inner pillar cells marked by p75 are present (Fig 8A and 8B), but pillar cells that are negative for S100A appear to be missing (Fig 5A). Since the inner pillar cell lies next to the inner hair cell, we speculate that outer pillar cell is more sensitive to the Fgf8 levels, and a decrease in the effective range of Fgf8 signaling due to reduced Fgf8 levels in the inner hair cells inSix1 CKO mutant might result in the lack of outer pillar cell formation. In addition to Fgf8, Six1-bound regions associated with Fgfr1 have also been identified by Six1 ChIP-seq
anal-ysis (J. Li, E. Loh, J. Xu, T. Zhang, L. Shen and P-X. Xu, manuscript in preparation). Given the similarity of the phenotype between theSix1 CKO and the Fgfr1 mutants, which range from
missing the outermost row of outer hair cells in mildly hypomorphicFgfr1 mutant to only
residual numbers of inner hair cells in the most severe mutants,Six1 may also regulate Fgfr1
signaling in the organ of Corti.
One intriguing finding of this study is the high level of Sox2 expression in theSix1 CKO hair
cells. While Sox2 is expressed in type II hair cells in the adult mouse utricle [46], it is absent from auditory hair cells. Sox2 is known to play a direct role in establishing the prosensory domain
within the cochlea, but Sox2 alone is unable to induceAtoh1 expression [8], as it interacts with Eya1 and Six1 to regulate the initiation ofAtoh1 expression [11]. However, following the onset of Atoh1 expression in the hair cell precursors, Sox2 levels become downregulated in differentiating hair cells, and an antagonistic interaction between Sox2 and Atoh1 was suggested to play a role in this downregulation [8]. InSix1 CKO cochlea, although the expression level of Atoh1 appears to
be lower than normal, it is sufficient to promote subsequent hair cell differentiation to generate Myo7a+hair cells even in the presence of high levels of Sox2. Thus, the capacity of endogenous Atoh1 to direct the hair cell differentiation program does not depend on its ability to downregu-late Sox2. In agreement with this view, forced expression of Atoh1 in cochlear epithelium in young mice is able to induce Sox2+cells to become ectopic Myo7a+hair cells in the GER [47], but Sox2 levels appear to drop in Myo7a+ectopic hair cells as their differentiation advances. Although detailed Six1 expression in postnatal cochlea has not been studied, the GER cells in young animals soon after birth are likely to retain some levels of Six1, which is widely expressed in the cochlear epithelium at birth (Fig 3). Our data show that in the absence of Six1, Sox2 does not appear to inhibit Atoh1 function in the differentiating hair cells. In the presence of Six1, deletion of Atoh1 in hair cells at later stages does not lead to upregulation of Sox2. Therefore, the antagonistic effect between Sox2 and Atoh1 is likely to be indirect and mediated through other factors, which may vary among different types of hair cells in the inner ear. This would explain why there are Sox2+ type II hair cells in the utricle and why a subset of Atoh1-induced high Sox2+ectopic hair cells exist in the cochlea [47]. While future studies are necessary to determine how many factors are involved in Sox2 downregulation in the auditory hair cells and how Six1 works together with them to repress Sox2 expression, our finding of high levels of Sox2 in Atoh1+hair cells inSix1
CKO cochlea uncovers a previously unknown function of Six1 in regulating the spatiotemporal pattern of Sox2 during the differentiation of the organ of Corti.
Our data show that Six1 is essential not only for hair cell fate induction, but also for proper patterning of the postmitotic precursor cells in the sensory epithelium. The precursor cells undergo rearrangements through both mediolateral and radial intercalation to achieve exten-sion and establish the mosaic structure between hair cells and supporting cells [4,48]. These processes require adhesion changes that allow cells to move and maintain adhesion, and cad-herins are known to control differential adhesive properties of cells during morphogenesis [49]. In the cochlea, the adhesion junction proteins E-cadherin and N-cadherin at the cell membrane mark a sharp boundary between the inner and outer hair cells and a direct involve-ment of these proteins in convergent extension in the cochlea has been shown recently [28]. InSix1 CKO cochlea, the sharp border formed by the expression of N-cadherin or E-cadherin
is disrupted and their expression is expanded to all cells in the organ of Corti. Such alterations in cadherin distribution are likely to lead to cellular morphological alterations. These data together support a role for Six1 in establishing the mosaic structure between hair cells and sup-porting cells in the organ of Corti.
Lastly, it is worth mentioning thatSix1 gene dosage may have differential effect on the
development between inner and outer hair cells, as the hypomorphicCwe/+ heterozygous
mice have extra inner hair cells in the apex of the cochlea, while their outer hair cells appear unaffected [14]. Similar phenotype also occurs inEya1+/-[50] andEya1CreERmice (Fig 6A). Thus, it is possible that Six1 may be required at a certain level at specific time points in cochlear development to regulate a different set of genes between inner and outer hair cell pre-cursors that are particularly sensitive to the level of Six1. Six1 may mediate organ of Corti for-mation through Fgf signaling, including Fgf8 signaling. While allelic series ofSix1 will provide
insight into its dosage effect on Fgf signaling and inner ear development, whole-genome map-ping of Six1-DNA interactions and identification of its direct targets at different developmen-tal stages will be necessary toward understanding how sensory progenitor cells use Six1 to
create precise patterns of gene expression and cell differentiation to shape and generate a func-tional organ of Corti for hearing in the mammalian inner ear.
Methods
Ethics statement
All animal protocols were approved by Animal Care and Use Committee of the Icahn School of Medicine at Mount Sinai (protocol #06–0807).
Animals, genotyping and tamoxifen administration
Six1Flox[17],Eya1CreERT2[11],Sox2CreERT2[19], andAtoh1Flox(JAX # 008681) mice were main-tained on a 129/Sv and C57BL/6J mixed background at the Icahn School of Medicine at Mount Sinai Animal Facility.
Mice were bred using timed mating, and noon on the day of vaginal plug detection was considered as E0.5. For induction of the CreERT2 protein, tamoxifen (Sigma, T5648) was dis-solved in corn oil (Sigma, C8267) and administrated (1.5 mg/10 g body weight) by oral gavage. Observed variations amongSix1 mutants is likely due to pre-existing developmental variation
between embryos when tamoxifen was given.
Histology, immunohistochemistry and in situ hybridization (ISH)
Histological examination, whole-mount and section immunostaining and ISH were carried out according to standard procedures. Briefly, inner ears were fixed in 4% paraformaldehyde (PFA) for 1 hr at 4˚C, dehydrated, and embedded in wax. Paraffin sections were generated at 6μm. For ISH, tissues were fixed overnight. We used five embryos for each genotype at each stage for each probe and the result was consistent in each embryo.
Primary antibodies: anti-Sox2 (PA1-094, Thermo Fisher), -Myo7A (25–6790, Proteus and 138-1-s, DSHB), -Six1 (HPA001893, Sigma), -Atoh1 (Math1-s, DSHB), -p27kip1(554069, BD Pharmingen), -Calretinin (MA5-14540, Thermo Fisher), -p75NTR(#07–476, EMD Millipore), -N-cadherin (610921, BD Bioscience), -E-cadherin (U3254, Sigma), -S100A (ab11428, Abcam), -GLAST (ab416, Abcam), -Pou4f3 (sc-81980, Santa Cruz), -Prox1 (AB5475, Milli-pore), -Acetylated tubulin (T7451, Sigma), -Cy3-, Cy2-, Cy5- and FITC-conjugated secondary antibodies were used. Alexa Fluor 488 or 350-conjugated phalloidin (A12379 and A22281, Life technologies) were used for actin staining. Hoechst 3342 was used for nuclear staining.
EdU and TUNEL assays
The EdU assay was performed using a kit (catalog no. C10640, Life Technologies) following the manufacturer’s instructions. EdU was co-injected with tamoxifen at 9 am of E11.5 and embryos were harvested at noon of E14.5. EdU was also injected at noon of E14.5 embryos fol-lowing tamoxifen treatment at 9 am of E11.5 and embryos were harvested at noon of E17.5. The TUNEL assay was performed using the Apop Tag kit for in situ apoptosis fluorescein detection (catalog no. NC9815837, Millipore) following the manufacturer’s instructions.
Reverse transcription and real-time PCR
Whole inner ears collected from E15.5 or E17.5 embryos were divided into two parts with for-ceps and the cochlear parts, which also contained the spiral ganglion, were used for total RNA extraction using Trizol Reagents (Invitrogen). Total RNAs were treated with RNase-Free DNase Set (QIAGEN) and then used for reverse transcription using a SuperScript IV Reverse Transcriptase (Thermo Fisher Scientific) for first-Strand cDNA Synthesis. Gene specific
primers and SYBR Green Master Mix (Applied Biosystems) were used for PCR amplification using the Applied Biosystems StepOnePlus Real-Time PCR Systems. Expression levels of each transcript were normalized usingβ-actin as an internal control. Each set of experiments was repeated three times, and the DDCT relative quantification method was used to evaluate quan-titative variation. Two-tailed Student’s t test was used for statistical analysis. Primers used are as follows: forAtoh1, forward primer-5’-GCTTATCCCCTTCGTTGAACT-3’ and reverse
primer-5’-TGCTATCCAGGAGGGACAGTTCTG-3’; forFgf8, forward primer-5’-ACGACAT
TCCACGAGCCGCGTC-3’ and reverse primer-5’-GAAGGGTCGGTCCTCGTGTCCCT-3’; and forβ-actin, forward primer 5’-AACGGCTCCGGCATGTGCAAAG-3’ and reverse primer 5’-ACACGCAGCTCATTGTAGAAG-3’.
Cell counts and spatial calibration
EdU-incorporated Sox2+prosensory progenitor cells in the floor of the cochlear epithelium were counted in basal, medial and apical turn of the entire cochlea. Values represent average number of EdU+Sox2+cells (±standard deviations) per section (6 μm) or per cochlea. Width and height of the Sox2+or p27kip1+domain at E14.5 were measured on sections (height at 6μm/section) for spatial calibration using Image J software (NIH). 15 sections per cochlea and 2 cochleae for each sample were measured. Two-tailed Student’s t test was used for statistical analysis.
Supporting information
S1 Fig. Six1 expression is lost in the hair cell precursors inSix1 CKO cochlea (given tamox-ifen at E11.5–12.5) using eitherEya1CreERT2orSox2CreERT2as a deletor. (A) Lineage tracing usingR26RLacZreporter confirmed that one dose of tamoxifen administration at E11.5 induced
Eya1CreERT2-lineage traced cells in the GER and all cells in the organ of Corti, including some Henson’s cells at P0. (B) Six1 (red)/p27Kip1 (green)costaining showing Six1 reduction in the sensory region inEya1CreER;Six1fl/fl. (C-E) Six1 (red) and Myo7a (green) section staining from E18.5 wild-type (C),Eya1CreERT2; Six1fl/fl(D) andSox2CreERT2; Six1fl/flcochlea (E). Scale bars: 30μm.
(PDF)
S2 Fig. Smaller inner ears and a slight delay in the development of the prosensory progeni-tors inSix1 CKO given tamoxifen at E11.5–12.5. (A,B) Six1 CKO mutants have smaller inner ears, compared toEya1CreERor wild-type littermates at E14.5 (A) and E17.5 (B). (C) Cochlear sections stained with p27Kip1(green) and EdU (red) from E17.5Eya1CreERorSix1
CKO littermate embryos injected with EdU at E14.5. Scale bar: 30μm. (PDF)
S3 Fig. Abnormal apoptosis was not significantly increased inSix1 CKO cochlea between E12.5-E14.5 given tamoxifen at E11.5-E12.5. TUNEL assay on sections from E14.5 wild-type (A,B) orSix1 CKO (Eya1CreERT2;Six1fl/fl) littermate cochlea (C,D) or E13.5Eya1CreER(E) or
Six1 CKO littermate cochlea (F) showing apoptotic cells in the cochlear epithelium (red). (G)
Statistical analysis showing average number of apoptotic cells in the floor of cochlear epithe-lium per section (6μm) at E12.5 (p = 0.023), E13.5 (p = 0.09) and E14.5 (p = 0.07). Scale bars: 100μm.
(PDF)
S4 Fig. Largely reduced utricular and saccular macula with fewer hair cells and no hair cells in crista ampullaris in all three semicircular canals. (A-F) Myo7a (green) and Sox2 (red) staining on sections of utricle (A,B), saccule (C,D) and crista (E,F) from E18.5 wild-type
orEya1CreERT2; Six1fl/flcochlea given tamoxifen at E11.5 and E12.5. Scale bars: 100μm. (PDF)
Acknowledgments
We thank J. Li, B.Y. Zhao, L. Zhang, and J.D. Labonne for technical assistance.
Author Contributions
Conceptualization: Ting Zhang, Pin-Xian Xu. Formal analysis: Ting Zhang, Jinshu Xu. Funding acquisition: Pin-Xian Xu.
Investigation: Ting Zhang, Jinshu Xu, Pin-Xian Xu. Project administration: Ting Zhang, Pin-Xian Xu. Resources: Pascal Maire.
Supervision: Pin-Xian Xu.
Validation: Ting Zhang, Jinshu Xu. Visualization: Ting Zhang.
Writing – original draft: Ting Zhang, Pin-Xian Xu.
Writing – review & editing: Ting Zhang, Pascal Maire, Pin-Xian Xu.
References
1. Cantos R, Cole LK, Acampora D, Simeone A, Wu DK. Patterning of the mammalian cochlea. Proc Natl Acad Sci U S A. 2000; 97(22):11707–13. Epub 2000/10/26.https://doi.org/10.1073/pnas.97.22.11707 PMID:11050199; PubMed Central PMCID: PMC34339.
2. Ruben RJ. Development of the inner ear of the mouse: a radioautographic study of terminal mitoses. Acta Otolaryngol. 1967:Suppl 220:1–44. Epub 1967/01/01. PMID:6067797.
3. Chen P, Segil N. p27(Kip1) links cell proliferation to morphogenesis in the developing organ of Corti. Development. 1999; 126(8):1581–90. Epub 1999/03/18. PMID:10079221.
4. Chen P, Johnson JE, Zoghbi HY, Segil N. The role of Math1 in inner ear development: Uncoupling the establishment of the sensory primordium from hair cell fate determination. Development. 2002; 129 (10):2495–505. PMID:11973280.
5. Wang J, Mark S, Zhang X, Qian D, Yoo SJ, Radde-Gallwitz K, et al. Regulation of polarized extension and planar cell polarity in the cochlea by the vertebrate PCP pathway. Nat Genet. 2005; 37(9):980–5. Epub 2005/08/24.https://doi.org/10.1038/ng1622PMID:16116426; PubMed Central PMCID: PMC1413588.
6. Kiernan AE, Pelling AL, Leung KK, Tang AS, Bell DM, Tease C, et al. Sox2 is required for sensory organ development in the mammalian inner ear. Nature. 2005; 434(7036):1031–5. Epub 2005/04/23. https://doi.org/10.1038/nature03487PMID:15846349.
7. Bermingham NA, Hassan BA, Price SD, Vollrath MA, Ben-Arie N, Eatock RA, et al. Math1: an essential gene for the generation of inner ear hair cells. Science. 1999; 284(5421):1837–41. Epub 1999/06/12. PMID:10364557.
8. Dabdoub A, Puligilla C, Jones JM, Fritzsch B, Cheah KS, Pevny LH, et al. Sox2 signaling in prosensory domain specification and subsequent hair cell differentiation in the developing cochlea. Proc Natl Acad Sci U S A. 2008; 105(47):18396–401. Epub 2008/11/18.https://doi.org/10.1073/pnas.0808175105 PMID:19011097; PubMed Central PMCID: PMC2587543.
9. Neves J, Parada C, Chamizo M, Giraldez F. Jagged 1 regulates the restriction of Sox2 expression in the developing chicken inner ear: a mechanism for sensory organ specification. Development. 2011; 138(4):735–44. Epub 2011/01/27.https://doi.org/10.1242/dev.060657PMID:21266409.
10. Neves J, Uchikawa M, Bigas A, Giraldez F. The prosensory function of Sox2 in the chicken inner ear relies on the direct regulation of Atoh1. PLoS One. 2012; 7(1):e30871.https://doi.org/10.1371/journal. pone.0030871PMID:22292066; PubMed Central PMCID: PMCPMC3264626.
11. Ahmed M, Wong EY, Sun J, Xu J, Wang F, Xu PX. Eya1-Six1 interaction is sufficient to induce hair cell fate in the cochlea by activating Atoh1 expression in cooperation with Sox2. Dev Cell. 2012; 22(2):377– 90.https://doi.org/10.1016/j.devcel.2011.12.006PMID:22340499; PubMed Central PMCID:
PMC3285434.
12. Zheng W, Huang L, Wei ZB, Silvius D, Tang B, Xu PX. The role of Six1 in mammalian auditory system development. Development. 2003; 130(17):3989–4000. Epub 2003/07/23. PMID:12874121. 13. Ozaki H, Nakamura K, Funahashi J, Ikeda K, Yamada G, Tokano H, et al. Six1 controls patterning of
the mouse otic vesicle. Development. 2004; 131(3):551–62. Epub 2003/12/26.https://doi.org/10.1242/ dev.00943PMID:14695375.
14. Bosman EA, Quint E, Fuchs H, Hrabe de Angelis M, Steel KP. Catweasel mice: a novel role for Six1 in sensory patch development and a model for branchio-oto-renal syndrome. Dev Biol. 2009; 328(2):285– 96. Epub 2009/04/25.https://doi.org/10.1016/j.ydbio.2009.01.030PMID:19389353; PubMed Central PMCID: PMC2682643.
15. Ruf RG, Xu PX, Silvius D, Otto EA, Beekmann F, Muerb UT, et al. SIX1 mutations cause branchio-oto-renal syndrome by disruption of EYA1-SIX1-DNA complexes. Proc Natl Acad Sci U S A. 2004; 101 (21):8090–5. Epub 2004/05/14.https://doi.org/10.1073/pnas.0308475101PMID:15141091; PubMed Central PMCID: PMC419562.
16. Konig R, Fuchs S, Dukiet C. Branchio-oto-renal (BOR) syndrome: variable expressivity in a five-genera-tion pedigree. Eur J Pediatr. 1994; 153(6):446–50. Epub 1994/06/01. PMID:8088301.
17. Le Grand F, Grifone R, Mourikis P, Houbron C, Gigaud C, Pujol J, et al. Six1 regulates stem cell repair potential and self-renewal during skeletal muscle regeneration. J Cell Biol. 2012; 198(5):815–32. https://doi.org/10.1083/jcb.201201050PMID:22945933; PubMed Central PMCID: PMCPMC3432771. 18. Xu J, Wong EY, Cheng C, Li J, Sharkar MT, Xu CY, et al. Eya1 interacts with Six2 and Myc to regulate
expansion of the nephron progenitor pool during nephrogenesis. Dev Cell. 2014; 31(4):434–47.https:// doi.org/10.1016/j.devcel.2014.10.015PMID:25458011; PubMed Central PMCID: PMC4282136. 19. Arnold K, Sarkar A, Yram MA, Polo JM, Bronson R, Sengupta S, et al. Sox2(+) adult stem and
progeni-tor cells are important for tissue regeneration and survival of mice. Cell Stem Cell. 2011; 9(4):317–29. https://doi.org/10.1016/j.stem.2011.09.001PMID:21982232; PubMed Central PMCID: PMC3538360. 20. Xu J, Ueno H, Xu CY, Chen B, Weissman IL, Xu PX. Identification of mouse cochlear progenitors that
develop hair and supporting cells in the organ of Corti. Nat Commun. 2017; 8:15046.https://doi.org/10. 1038/ncomms15046PMID:28492243.
21. Gu R, Brown RM, 2nd, Hsu CW, Cai T, Crowder AL, Piazza VG, et al. Lineage tracing of Sox2-express-ing progenitor cells in the mouse inner ear reveals a broad contribution to non-sensory tissues and insights into the origin of the organ of Corti. Dev Biol. 2016; 414(1):72–84.https://doi.org/10.1016/j. ydbio.2016.03.027PMID:27090805; PubMed Central PMCID: PMCPMC4875846.
22. Chonko KT, Jahan I, Stone J, Wright MC, Fujiyama T, Hoshino M, et al. Atoh1 directs hair cell differenti-ation and survival in the late embryonic mouse inner ear. Dev Biol. 2013; 381(2):401–10.https://doi.org/ 10.1016/j.ydbio.2013.06.022PMID:23796904; PubMed Central PMCID: PMCPMC3772529.
23. Kiernan AE, Xu J, Gridley T. The Notch ligand JAG1 is required for sensory progenitor development in the mammalian inner ear. PLoS Genet. 2006; 2(1):e4.https://doi.org/10.1371/journal.pgen.0020004 PMID:16410827; PubMed Central PMCID: PMCPMC1326221.
24. Jones JM, Montcouquiol M, Dabdoub A, Woods C, Kelley MW. Inhibitors of differentiation and DNA binding (Ids) regulate Math1 and hair cell formation during the development of the organ of Corti. J Neu-rosci. 2006; 26(2):550–8.https://doi.org/10.1523/JNEUROSCI.3859-05.2006PMID:16407553. 25. Jacques BE, Montcouquiol ME, Layman EM, Lewandoski M, Kelley MW. Fgf8 induces pillar cell fate
and regulates cellular patterning in the mammalian cochlea. Development. 2007; 134(16):3021–9. https://doi.org/10.1242/dev.02874PMID:17634195.
26. Jones C, Chen P. Primary cilia in planar cell polarity regulation of the inner ear. Curr Top Dev Biol. 2008; 85:197–224. Epub 2009/01/17.https://doi.org/10.1016/S0070-2153(08)00808-9PMID: 19147007.
27. Heisenberg CP, Bellaiche Y. Forces in tissue morphogenesis and patterning. Cell. 2013; 153(5):948– 62.https://doi.org/10.1016/j.cell.2013.05.008PMID:23706734.
28. Chacon-Heszele MF, Ren D, Reynolds AB, Chi F, Chen P. Regulation of cochlear convergent extension by the vertebrate planar cell polarity pathway is dependent on p120-catenin. Development. 2012; 139 (5):968–78.https://doi.org/10.1242/dev.065326PMID:22318628; PubMed Central PMCID: PMCPMC3274358.
29. Li J, Rodriguez Y, Cheng C, Zeng L, Wong EY, Xu CY, et al. EYA1’s Conformation Specificity in Dephosphorylating Phosphothreonine in Myc and Its Activity on Myc Stabilization in Breast Cancer. Mol Cell Biol. 2017; 37(1).https://doi.org/10.1128/MCB.00499-16PMID:27795300; PubMed Central PMCID: PMCPMC5192086.
30. Puligilla C, Kelley MW. Dual role for Sox2 in specification of sensory competence and regulation of Atoh1 function. Dev Neurobiol. 2016.https://doi.org/10.1002/dneu.22401PMID:27203669.
31. Munnamalai V, Fekete DM. Notch-Wnt-Bmp crosstalk regulates radial patterning in the mouse cochlea in a spatiotemporal manner. Development. 2016; 143(21):4003–15.https://doi.org/10.1242/dev. 139469PMID:27633988; PubMed Central PMCID: PMCPMC5117145.
32. Ozaki H, Watanabe Y, Takahashi K, Kitamura K, Tanaka A, Urase K, et al. Six4, a putative myogenin gene regulator, is not essential for mouse embryonal development. Mol Cell Biol. 2001; 21(10):3343– 50. Epub 2001/04/21.https://doi.org/10.1128/MCB.21.10.3343-3350.2001PMID:11313460; PubMed Central PMCID: PMC100256.
33. Schlosser G. Induction and specification of cranial placodes. Dev Biol. 2006; 294(2):303–51. Epub 2006/05/09.https://doi.org/10.1016/j.ydbio.2006.03.009PMID:16677629.
34. Konishi Y, Ikeda K, Iwakura Y, Kawakami K. Six1 and Six4 promote survival of sensory neurons during early trigeminal gangliogenesis. Brain Res. 2006; 1116(1):93–102.https://doi.org/10.1016/j.brainres. 2006.07.103PMID:16938278.
35. Suzuki Y, Ikeda K, Kawakami K. Expression of Six1 and Six4 in mouse taste buds. J Mol Histol. 2010; 41(4–5):205–14.https://doi.org/10.1007/s10735-010-9280-8PMID:20668922.
36. Ando Z, Sato S, Ikeda K, Kawakami K. Slc12a2 is a direct target of two closely related
homeobox proteins, Six1 and Six4. FEBS J. 2005; 272(12):3026–41. https://doi.org/10.1111/j.1742-4658.2005.04716.xPMID:15955062.
37. Zou D, Silvius D, Davenport J, Grifone R, Maire P, Xu PX. Patterning of the third pharyngeal pouch into thymus/parathyroid by Six and Eya1. Dev Biol. 2006; 293(2):499–512. Epub 2006/03/15.https://doi. org/10.1016/j.ydbio.2005.12.015PMID:16530750.
38. Streit A. Extensive cell movements accompany formation of the otic placode. Dev Biol. 2002; 249 (2):237–54. Epub 2002/09/11. PMID:12221004.
39. Grifone R, Demignon J, Houbron C, Souil E, Niro C, Seller MJ, et al. Six1 and Six4 homeoproteins are required for Pax3 and Mrf expression during myogenesis in the mouse embryo. Development. 2005; 132(9):2235–49. Epub 2005/03/25.https://doi.org/10.1242/dev.01773PMID:15788460.
40. Visel A, Thaller C, Eichele G. GenePaint.org: an atlas of gene expression patterns in the mouse embryo. Nucleic Acids Res. 2004; 32(Database issue):D552–6.https://doi.org/10.1093/nar/gkh029 PMID:14681479; PubMed Central PMCID: PMCPMC308763.
41. Hoskins BE, Cramer CH, Silvius D, Zou D, Raymond RM, Orten DJ, et al. Transcription factor SIX5 is mutated in patients with branchio-oto-renal syndrome. Am J Hum Genet. 2007; 80(4):800–4. Epub 2007/03/16.https://doi.org/10.1086/513322PMID:17357085; PubMed Central PMCID: PMC1852719. 42. Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, et al. A human homologue of the
Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat Genet. 1997; 15(2):157–64. Epub 1997/02/01.https://doi.org/10.1038/ng0297-157PMID: 9020840.
43. Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, et al. Clustering of mutations responsible for branchio-oto-renal (BOR) syndrome in the eyes absent homologous region (eyaHR) of EYA1. Hum Mol Genet. 1998/03/21 ed1997. p. 2247–55.
44. Shim K, Minowada G, Coling DE, Martin GR. Sprouty2, a mouse deafness gene, regulates cell fate decisions in the auditory sensory epithelium by antagonizing FGF signaling. Dev Cell. 2005; 8(4):553– 64.https://doi.org/10.1016/j.devcel.2005.02.009PMID:15809037.
45. Yousaf R, Meng Q, Hufnagel RB, Xia Y, Puligilla C, Ahmed ZM, et al. MAP3K1 function is essential for cytoarchitecture of the mouse organ of Corti and survival of auditory hair cells. Dis Model Mech. 2015; 8 (12):1543–53.https://doi.org/10.1242/dmm.023077PMID:26496772; PubMed Central PMCID: PMCPMC4728323.
46. Oesterle EC, Campbell S, Taylor RR, Forge A, Hume CR. Sox2 and JAGGED1 expression in normal and drug-damaged adult mouse inner ear. J Assoc Res Otolaryngol. 2008; 9(1):65–89. Epub 2007/12/ 25.https://doi.org/10.1007/s10162-007-0106-7PMID:18157569; PubMed Central PMCID:
PMC2536811.
47. Kelly MC, Chang Q, Pan A, Lin X, Chen P. Atoh1 directs the formation of sensory mosaics and induces cell proliferation in the postnatal mammalian cochlea in vivo. J Neurosci. 2012; 32(19):6699–710. https://doi.org/10.1523/JNEUROSCI.5420-11.2012PMID:22573692; PubMed Central PMCID: PMCPMC3477623.
48. McKenzie E, Krupin A, Kelley MW. Cellular growth and rearrangement during the development of the mammalian organ of Corti. Dev Dyn. 2004; 229(4):802–12.https://doi.org/10.1002/dvdy.10500PMID: 15042704.
49. Steinberg MS. Differential adhesion in morphogenesis: a modern view. Curr Opin Genet Dev. 2007; 17 (4):281–6.https://doi.org/10.1016/j.gde.2007.05.002PMID:17624758.
50. Zou D, Erickson C, Kim EH, Jin D, Fritzsch B, Xu PX. Eya1 gene dosage critically affects the develop-ment of sensory epithelia in the mammalian inner ear. Hum Mol Genet. 2008; 17(21):3340–56. Epub 2008/08/06.https://doi.org/10.1093/hmg/ddn229PMID:18678597.