HAL Id: tel-01233446
https://tel.archives-ouvertes.fr/tel-01233446
Submitted on 25 Nov 2015HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
termes de services écosystémiques
Stéphane Guitet
To cite this version:
Stéphane Guitet. Diversité des écosystèmes forestiers de Guyane française : distribution, détermi-nants et conséquences en termes de services écosystémiques. Biodiversité et Ecologie. Université de Montpellier, 2015. Français. �tel-01233446�
Délivré par l’Université de Montpellier
Préparée au sein de l’école doctorale SIBAGHE
Et de l’unité de recherche UMR AMAP
Spécialité
Spécialité
Spécialité
Spécialité : Ecosystèmes et Sciences Agronomiques
: Ecosystèmes et Sciences Agronomiques
: Ecosystèmes et Sciences Agronomiques
: Ecosystèmes et Sciences Agronomiques
Présentée par Stéphane GUITET
Soutenue le 25 Septembre 2015 devant le jury composé de
Jean-François DHOTE, DR, INRA (EFPA – Val de Loire) Rapporteur
Michel GRIMALDI, DR, IRD (iEES - Paris) Rapporteur
Hans TER STEEGE, Professeur, (NBC – Pays-Bas) Examinateur Pierre COUTERON, DR, IRD (AMAP - Montpellier) Directeur Daniel SABATIER, CR1, IRD (AMAP - Montpellier) Co-Directeur Thibaut DECAENS, Professeur,UM (CEFE) Président du Jury
Diversité des écosystèmes forestiers de
Guyane française : distribution,
déterminants et conséquences en termes de
services écosystémiques
1
Ces travaux sont dédicacés à ma tribu : Cécile, Maxime, Jérémie, Mathias …
beaucoup délaissés ces derniers mois mais toujours autant aimés
2
et ami, forestier passionné par la Guyane et fin botaniste, dont la contribution a été essentielle pour rassembler le jeu de données exceptionnel sur lequel j’ai pu appuyer ces travaux. Pour ton investissement, ta constance et les encouragements que tu m’as apportés au cours de ces dernières années soit mille fois remercié. Si j’ai pu réaliser cette thèse c’est aussi grâce à l’attention de Pierre Couteron qui, en tant que Directeur d’Unité, m’a tout d’abord accueilli à l’UMR Amap, puis a accepté de jouer le rôle de Directeur de thèse, me faisant ainsi bénéficier de sa vision éclairée sur des sujets ardus. Les conseils que Daniel Sabatier, mon co-directeur, et Raphaël Pélissier m’ont prodigué tout au long de ces travaux et la disponibilité dont ils ont fait preuve m’ont été extrêmement précieux pour orienter mes réflexions et améliorer mes connaissances. Pierre, Daniel et Raphaël, merci de tout cœur.
L’aide de Vincent Freycon, de l’UR BESF au CIRAD, a aussi été décisive et m’a permis d’aborder un volet qui me tenait particulièrement à cœur : la pédologie. C’est Vincent qui m’a guidé sur le tortueux chemin du WRB !... Merci à toi Vincent.
Je suis aussi redevable à de nombreux collègues avec qui j’ai collaboré, sur le terrain en Guyane et dans les labos à Montpellier, pour mener à bien cette thèse :
- Mes compagnons de forêts : Cécile Richard-Hansen, Sophie Gonzalez, Jean-Pierre Simonet, Vincent Bezard, les nombreux prospecteurs de l’ONF notamment Atidong Nano qui nous a accompagné sur tous les terrains, ainsi que les personnels du Parc Amazonien notamment François Bagadi et bien d’autres…
- Les personnes qui m’ont conseillé, aidé, ou accompagné, un jour ou toujours : Gaëlle Viennois, Gaëlle Jaouen, Bruno Hérault, Jean-Jacques de Granville, Christopher Baraloto, Maël Dewynter, Pierre Joubert, Thomas Denis, Maxime Réjou-Méchain, Nicolas Barbier, François Munoz, Jean-François Molino, Grégoire Vincent, Lilian Blanc, Martial Bernoux, Quentin Molto, Valéry Gond – la liste n’est pas exhaustive… L’AMAP n’a pas été seulement un laboratoire mais aussi une terre d’accueil. Merci à tous ceux qui m’ont aidé à supporter ce long séjour professionnel « hors sol » grâce à des poses méridiennes sympathiques. Merci aux coureurs Stéphanie, François, Christophe et le regretté Mickaël. Merci aux Paléos qui m’ont adopté : Jean, Brigitte, Anne-Laure. Aux efficaces « supportrices »: Noémie, Valérie, Yannick, Marie-Hélène, Nathalie. Merci à Jérôme, Gilles, Vincent, Cécile, Jean-Luc, Julien et tous les autres sympathiques affamés.
Enfin, merci à mon employeur l’Office National des Forêts et aux directeurs qui m’ont encouragé à me lancer dans cette odyssée principalement Jean-François Dhôte (ONF), Eric Marcon (ECOFOG), Daniel Auclair (INRA).
3
biodiversité, un bon état de conservation, une forte contribution au stock de carbone amazonien et une dynamique socio-économique très rapide. Les connaissances sur la diversité des écosystèmes forestiers de Guyane française restent cependant insuffisantes pour asseoir une gestion optimale des différents services écosystémiques à l’échelle du territoire. Les travaux réalisés dans le cadre de cette thèse, visent prioritairement à étudier la diversité des écosystèmes forestiers de Guyane, les facteurs déterminant leur variabilité et les conséquences de cette variabilité en termes de services écosystémiques et de gestion.
La première partie de la thèse met en évidence les différentes échelles de structuration spatiale de la diversité forestière et les patrons de composition qui en résultent à l’échelle régionale. Elle s’appuie sur une récente campagne d’inventaires forestiers (2006-2013) couvrant l’ensemble du département et rassemblant 3 132 placettes de 0.2ha implantées selon un plan d’échantillonnage emboîté et pluristratifié. Les résultats obtenus vérifient l’hypothèse d’une forte influence de la géomorphologie, à l’échelle des paysages, sur la bêta -diversité des forêts guyanaises en termes d’espèces dominantes (métrique de Simpson). Cette première étape a nécessité plusieurs développements méthodologiques préalables notamment l’évaluation de l’incertitude taxonomique des inventaires forestiers et sa propagation dans les analyses de diversité, ainsi que l’élaboration de modèles cartographiques permettant de caractériser précisément l’environnement géomorphologique à diverses échelles d’études.
La deuxième partie de la thèse vise à expliquer l’influence exercée par les paysages géomorphologiques sur la végétation forestière à travers l’étude des relations entre géomorphologie et sols. Elle s’appuie sur l’analyse de plus de 400 sondages pédologiques couplés à la campagne d’inventaires forestiers. Les résultats obtenus démontrent une forte dépendance de la nature et des propriétés des sols vis-à-vis des paysages géomorphologiques et du dénivelé régional. Ils suggèrent l’existence d’un effet de filtre édaphique sous-jacent au déterminisme géomorphologique sur la diversité forestière. Une part importante de l’influence géomorphologique reste cependant indépendante du filtre édaphique et est interprétée comme un effet conjugué de deux autres mécanismes sur deux échelles de temps distinctes : le contrôle du turn-over forestier par la dynamique géomorphologique au cours du quaternaire récent et l’intégration des effets biogéographiques sous l’influence des changements globaux plus anciens.
La troisième partie de la thèse s’intéresse aux conséquences de la diversité biotique et abiotique décrite sur la variabilité des services écosystémiques rendus par la
4
service de régulation est mis en relation avec la richesse spécifique locale (service de support) et la quantité de bois exploitable (service d’approvisionnement). Des relations complexes de corrélation et de compromis sont mises en évidence entre ces différents services. Elles illustrent la variabilité des réponses des services écosystémiques à la diversité des écosystèmes forestières.
La mise en perspective de ces différents volets permet de proposer une typologie précise des habitats forestiers de Guyane et de fournir des pistes de prise en compte de cette diversité écosystémique pour l’aménagement du territoire et la gestion forestière.
Mots-clefs : Ecologie des communautés, Forêt tropicale humide, Guyane française, Gestion forestière, Habitats naturels
5
Diversity of ecosystems in French Guiana rainforest: spatial
distribution, drivers and implications for ecosystems services and
forest management
French Guiana forest presents a high originality in terms of biodiversity, a good state of preservation, a strong contribution to the overall Amazonian carbon stocks and a very fast socio-economic dynamic. However, our knowledge about forest ecosystems diversity remains insufficient to ensure an optimal management of the various ecosystems and of the services they may provide across the territory. This thesis aims to answer this central issue by studying the diversity of forest ecosystems at the scale of the entire forested area of French Guiana. Specifically we targeted highlighting the factors that determine ecosystems variability and evaluating the consequences of this variability in terms of ecosystem services and management. Firstly, we bring out complex patterns of diversity at different scales including strong floristic variation at the regional scale, using a recent forest inventories campaign (2006-2013) that has been conducted according to a stratified sampling design which includes 3,132 0.2ha-plots covering the whole territory. Our results verify the hypothesis of a strong control of forest beta and alpha diversity by geomorphology, especially at the landscape scale and for dominant species (Simpson metric). This first step required several preliminary methodological developments including an assessment of the taxonomic uncertainty in forest inventories, and spatial models to accurately characterize the geomorphological context at various scales.
Secondly, we aimed at interpreting the influence of geomorphological landscapes on the composition of forest vegetation by exploring the relationship between geomorphology and soils. Using more than 400 soil sampling locations coupled with the forest inventory plots, we demonstrated that nature and properties of soils are dependent on geomorphological landscapes and locally distributed along regional elevation gradients. A significant soil filtering effect was found underlying the geomorphological influence on forest composition and diversity. However, a major part of this influence proved to be independent of soil filtering and is interpreted as a combination of two mechanisms at two different time-scales: (i) the control of forest turn-over by the geomorphological dynamics during the Late Quaternary and (ii) the integration of biogeographic effects under the influence of tectonic evolution and ancient climate changes.
Last of all, we addressed the consequences of these biotic and abiotic variations, which combine into habitats diversity, on forest management and on the variability
6
regulation service is related with the local species richness (pertaining to the category of support services) and the amount of usable wood (pertaining to the category of supply services). Complex relationships, including correlations and compromises, are evidenced between these different services. This illustrates the variability of the responses of ecosystem services to the habitats diversity.
Finally our work allowed us to provide a new typology of natural habitats present in French Guiana forest and to propose suggestions in order to improve land uses and multifunctional forest management in French Guiana.
Key-words : Community ecology, tropical rainforest, French Guiana, Forest management, Natural habitat
7
I Introduction générale ... 11
I.1 La gestion des forêts tropicales humides, cadre thématique de la thèse ... 11
I.1.a Les forêts tropicales humides au cœur des grands enjeux sociétaux .... 11
I.1.b La gestion durable multifonctionnelle ... 13
I.1.c La caractérisation des écosystèmes ... 15
I.2 La Guyane française et son cadre régional ... 17
I.2.a Le Bouclier guyanais sous-région amazonienne ... 17
I.2.b La diversité forestière de la sous-région guyanaise ... 18
I.2.c Contexte socio-économique en Guyane française ... 20
I.3 Cadre scientifique et structure de la thèse ... 22
I.3.a Objectifs et questions de recherche ... 22
I.3.b Cadre méthodologique ... 23
I.3.c Structure du document ... 28
II Diversité et distribution ... 30
II.1 Introduction au chapitre ... 30
II.1.a Etat des connaissances sur la diversité forestière guyanaise ... 30
II.1.b La géomorphologie indicatrice de la diversité forestière ? ... 35
II.1.c Travaux préalables à une étude régionale de la diversité ... 36
II.2 Mesure de la diversité régionale à partir d’inventaires forestiers ... 39
II.3 Etude des patrons de bêta-diversité dans un cadre multi-échelle ... 40
II.4 Synthèse du chapitre ... 41
III Déterminants et mécanismes ... 44
III.1 Introduction au chapitre ... 44
III.1.a Hypothèses relatives à l’influence de la géomorphologie sur la diversité : ... 45
III.1.b Faire la part des effets de niche... 48
8
III.4.a Un effet niche limité par des gradients environnementaux courts ... 52
III.4.b Une lecture dynamique des paysages géomorphologiques ... 54
IV Diversité et Services écosystémiques ... 59
IV.1 Introduction au chapitre ... 59
IV.1.a Définition des services écosystémiques ... 59
IV.1.b Régulation du cycle du carbone et approvisionnement en bois ... 60
IV.2 Modèle de distribution spatiale de la biomasse forestière à l’échelle régionale ... 62
IV.3 Variabilité régionale des SE et relations entre services ... 63
IV.3.a Matériels & méthodes ... 63
IV.3.b Résultats ... 66
IV.4 Synthèse du chapitre ... 72
V Conclusion générale ... 73
V.1 Une nouvelle classification des habitats forestiers de Guyane ... 73
V.2 Implications en termes de gestion forestière ... 76
V.2.a Une stratégie d’acquisition des connaissances affinée : ... 76
V.2.b Une stratégie de conservation axée vers la représentativité: ... 77
V.2.c Une aide à l’aménagement du Domaine Forestier Permanent (DFP) : .. 78
V.3 Limites et perspectives de ces travaux ... 79
V.4 Le mot de la fin ... 80
VI Références bibliographiques ... 81
9
FIGURE 1 : CONTRIBUTION RELATIVE DES FTH AUX PRINCIPAUX ENJEUX SOCIETAUX ET ENVIRONNEMENTAUX... 12 FIGURE 2 : REPRESENTATION DES INTERACTIONS ENTRE LES ECHELLES ECOLOGIQUES ET INSTITUTIONNELLES POUR DIFFERENTS SERVICES ECOSYSTEMIQUES ... 14 FIGURE 3(TIREE DE [60]) :CARTOGRAPHIE DES PRINCIPALES REGIONS FORESTIERES DU BOUCLIER DES GUYANES D’APRES TER STEEGE &ZONDERVAN ... 19 FIGURE 4 (TIREE DE [62]) : CARTOGRAPHIE DES TYPES DE PAYSAGES FORESTIERS DE GOND REALISEE PAR TELEDETECTION ... 19 FIGURE 5 :CARTOGRAPHIE DES ZONES FORESTIERES REGLEMENTAIRES EN VIGUEUR EN GUYANE .. 21 FIGURE 6 :SCHEMA D’ARTICULATION DE LA THESE. ... 29 FIGURE 7 : CARTOGRAPHIE DES SITES D’ETUDES DE LA DIVERSITE FORESTIERE MIS EN PLACE EN GUYANE DEPUIS LES ANNEES 1970 ... 31 FIGURE 8 :CARTE DES PAYSAGES GEOMORPHOLOGIQUES TIREE DE [126] ISSUE D’UNE ANALYSE DU MODELE D’ELEVATION DU SRTM-30M ... 37 FIGURE 9 : DISPOSITIF D’ECHANTILLONNAGE DE L’INVENTAIRE HABITATS EXPLOITE DANS LE CADRE DE LA THESE. ... 38 FIGURE 10 : FORTE STRUCTURATION DE LA DIVERSITE A LARGE ECHELLE ET ABSENCE D’ORGANISATION NETTE AUX ECHELLES INFERIEURES DEMONTREES PAR SCALOGRAMMES SELON LES METHODES PROPOSEES PAR [68]. ... 42 FIGURE 11 :MODELE DE DISTRIBUTION D’ABONDANCE POUR DEUX ESPECES D’ARBRES GUYANAIS UTILISANT UNE FONCTION LOESS DU SECOND DEGRE SELON LA METHODE PROPOSEE PAR [42] BASEE SUR LES POSITIONS GEOGRAPHIQUES A RESOLUTION DE 0,125°(ENVIRON 14KM) ... 43 FIGURE 12 (TIREE DE [130]):REPRESENTATION DES QUATRE GRANDS PROCESSUS IMPLIQUES DANS LES MECANISMES DE DIVERSIFICATION DES ECOSYSTEMES COMPLEXES ... 44 FIGURE 13 (TIREE DE [144]):SCHEMA REPRESENTATIF DES DIVERGENCES DE DYNAMIQUE ENTRE LES ECOSYSTEMES AMORTIS (DAMPENING SYSTEM) ET ECOSYSTEMES AGRESSIFS (RAMPING SYSTEM) ET LEUR PRINCIPALES CARACTERISTIQUES BIOTIQUES ET ABIOTIQUES ... 46 FIGURE 14 : SCHEMA DES RELATIONS SUPPOSEES ENTRE PAYSAGES GEOMORPHOLOGIQUES ET DIVERSITE FORESTIERE DANS LE CONTEXTE GUYANAIS. ... 49 FIGURE 15 : ANALYSE EN COMPOSANTES PRINCIPALES DES FACTEURS PEDOLOGIQUES ET CLIMATIQUES. ... 52
10
FIGURE 17 :REPRESENTATION SIMPLIFIEE DU MODELE DE DISTRIBUTION DES TYPES DE SOLS WRB ... 54 FIGURE 18 :POSITIONNEMENT DES FOSSES PEDOLOGIQUES OUVERTES SUR LE SITE DE LA WAKI LE LONG D’UNE TOPO SEQUENCE. ... 56 FIGURE 19 : EVOLUTION DU RAPPORT ISOTOPIQUE DU C13 SUR LE PROFIL F1 DE LA WAKI ET
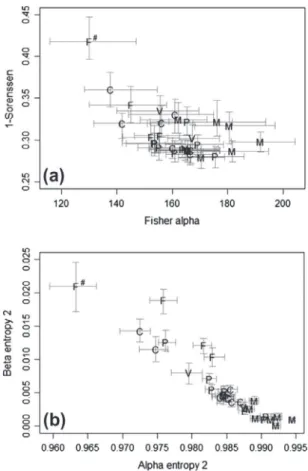
COMPARAISON AVEC UN PROFIL DE PARACOU (TYPIQUE D’UN COUVERT FORESTIER PERMANENT).. 57 FIGURE 20 :COMPARAISON DES VALEURS DE FISHER-ALPHA ESTIMEES A L’ECHELLE DES SITES A PARTIR DES INVENTAIRES FORESTIERS... 58 FIGURE 21 :PROJECTION DES COORDONNEES DES LAYONS SUR LES 2 PREMIERS AXES DE L’ANALYSE EN COMPOSANTE PRINCIPALE DES 10 INDICATEURS DE SE ... 67 FIGURE 22 :RELATION ENTRE STOCKS DE CARBONE ET DIVERSITE ALPHA DE FISHER PAR SITES ET FONCTIONS DE REGRESSION ASSOCIEES ... 69 FIGURE 23 :STOCK DE CARBONE (BIOMASSE AERIENNE ET SOL) EN FONCTION DE LA DENSITE DES TIGES EXPLOITABLES D’ESSENCES COMMERCIALES PAR LAYONS ET FONCTION DE REGRESSION ASSOCIEE EN LOG-LOG. ... 69 FIGURE 24 : RELATION ENTRE DIVERSITE LOCALE ET DENSITE DES ESSENCES COMMERCIALES PRINCIPALES (DBH≥20CM) PAR LAYONS ET FONCTION DE REGRESSION ASSOCIEE ... 71 FIGURE 25 :RELATION ENTRE L’INDICE DE DIVERSITE ALPHA BASE SUR L’ENTROPIE D’ORDRE 2 (SIMPSON) ET LA DENSITE EN TIGES EXPLOITABLES D’ESSENCES COMMERCIALES MAJEURES (DBH≥55CM) PAR SITES. ... 71 FIGURE 26 :EFFORT DE PROTECTION PAR TYPE DE PAYSAGE ... 77
11
Chapitre Introductif
I
Introduction générale
I.1
La gestion des forêts tropicales humides, cadre
thématique de la thèse
I.1.a
Les forêts tropicales humides au cœur des grands enjeux
sociétaux :
alimentation,
biodiversité,
climat,
développement
De par leur vaste étendue, la diversité biologique qu’elles renferment, la concentration de biomasse qu’elles représentent et la place qu’elles occupent dans les cycles biogéochimiques globaux, les forêts tropicales humides (FTH) sont au cœur d’importants enjeux écologiques qui conditionnent le devenir de notre planète et l’avenir de nos sociétés (Figure 1).
Le premier enjeu concernant les FTH est celui de l’alimentation des 9 milliards d’êtres humains qui peupleront demain la planète [1,2]. Cet impératif se traduit aujourd’hui par la conversion depuis une vingtaine d’année de plus de 6 millions d’hectares de FTH par an, soit en plantations de palmiers à huile (près de 8 millions d’hectares aujourd’hui en Indonésie et Malaisie [3]), en pâturages ou en cultures de soja (plus de 56 millions d’hectares en Amazonie [4]). Cette déforestation rapide évaluée à -0.39% par an [5] répondant à l’approvisionnement du marché agro-alimentaire mondial constitue une véritable menace pour la biodiversité abritée par ces écosystèmes qui représenterait 50 à 60% de toutes les espèces vivantes actuelles [6].
Bien que seulement 3 des 25 hot-spots de biodiversité [7] concernent les grands bassins forestiers tropicaux, ceux-ci regroupent plus de 40% des espèces végétales menacées [8], 29 des 200 écorégions de conservation prioritaires reconnues pour leur valeur représentative ou leur taux d’endémisme [9] et 3 des 5 grandes régions sauvages à forte biodiversité [10].
La pression qui s’exerce sur les FTH ne menace pas seulement la richesse du patrimoine naturel mondial mais plus largement les conditions de son existence. La destruction des FTH est en effet la source de profondes modifications climatiques régionales par l’augmentation des températures et la réduction des précipitations locales [4]. Mais elle participe également aux dérèglements des grands cycles biogéochimiques qui conditionnent les changements climatiques en cours, en étant
12
responsable de 15 à 20% de l’émission mondiale des gaz à effets de serre et en premier lieu du relargage net annuel d’environ 1 milliard de tonnes de carbone jusque-là immobilisé dans la biomasse forestière [11]. Ce faisant, la déforestation contribue aussi à réduire l’important effet puits de carbone produit par l’activité photosynthétique de ce capital forestier (grossièrement estimée à 1.2 GtC.an-1
[12,13]) ce qui amplifie l’accumulation du CO2 dans l’atmosphère.
FIGURE 1 : CONTRIBUTION RELATIVE DES FTH AUX PRINCIPAUX ENJEUX SOCIETAUX ET ENVIRONNEMENTAUX
Les chiffres sont tirés des statistiques nationales des 30 pays appartenant aux trois grands bassins forestiers tropicaux définis par la FAO [5] couverts par la FTH pour les deux tiers de leur surface. Les chiffres concernant les espèces menacées ont été calculés à partir des RedList de l’IUCN [8]
Parallèlement, les bassins forestiers tropicaux, longtemps peu peuplés, suivent une dynamique démographique rapide (+1.4% par an) et abritent aujourd’hui une population forte de plus d’un milliard d’êtres humains [5]. De fait, l’exploitation des ressources forestières (provenant du bois, des sols, ou du sous-sol) ne cesse de s’accélérer.
Concilier le développement économique, et notamment l’activité agricole vitale pour les populations locales, et la préservation de l’intégrité de ces écosystèmes, essentielle à l’échelle planétaire, est par conséquent un des principaux défis qui s’imposent aujourd’hui aux instances politiques, citoyennes et scientifiques en charge d’orienter les décisions relatives à l’aménagement de ces territoires.
13
I.1.b
La gestion durable multifonctionnelle : une réponse à la
multiplicité des enjeux ?
Comment concilier ces enjeux apparemment antagonistes dans des écosystèmes aussi complexes que les forêts tropicales humides ? Les solutions proposées pour relever ce défi font appel à deux notions fondamentales qui ont émergées il y a une vingtaine d’année de cela : la notion de « gestion et développement durable » (GD) et celles de « services écosystémiques » (SE).
Le concept de GD, mis en avant en 1992 lors de la désormais célèbre conférence de la CNUED (Conférence des Nations Unies sur l’Environnement et le Développement) à Rio, définit comme objectif la valorisation des écosystèmes de manière à maintenir leur fonctionnement biologique (productivité, vitalité, capacité de régénération, diversité) et leur capacité à fournir les mêmes niveaux de biens et services aux générations à venir [14]. La notion de SE, qui émerge à la même période [15] mais ne sera massivement développée que 10 ans plus tard [16], vient préciser et renforcer la notion de durabilité en pointant le rôle essentiel des fonctions de supports des écosystèmes (biodiversité, sols, production primaire…) dans la fourniture des biens et services attendus par la société. Le concept de gestion durable et multifonctionnelle (c’est-à-dire garantissant l’équilibre entre les différents SE) qui s’en dégage est loin d’être nouveau dans le monde forestier tempéré où les notions de planification et de long terme sont déjà répandues depuis plusieurs siècles, notamment en Europe [17]. La nouveauté réside dans sa généralisation à la forêt tropicale et dans l’élargissement de cette notion de durabilité à une sphère beaucoup plus large que la seule gestion forestière : celle du développement dans son ensemble et de l’aménagement du territoire.
Cependant la transposition de ce concept aux FTH se heurte à une double difficulté : celle relative à la complexité de ces écosystèmes hyper-diversifiés et celle relative à la multiplication des échelles concernées. Si la régulation des grands cycles biogéochimiques nécessite en effet une approche globale aussi bien pour la compréhension des mécanismes que pour les prises de décisions internationales, les stratégies de conservation de la biodiversité doivent quant à elles plutôt être étudiées et optimisées à l’échelle d’écorégions [9]. Mais pour la plupart des SE (comme la gestion des eaux de surface, la maîtrise de l’érosion, l’approvisionnement en ressources) c’est aux échelles intermédiaires (paysage, écosystème, parcelle – Figure 2) qu’opèrent les mécanismes [18]. L’acquisition de connaissances sur les fonctionnements écosystémiques aux échelles intermédiaires est donc essentielle. Elle est d’autant plus nécessaire que les mécanismes écologiques et économiques régissant ces systèmes complexes sont très dépendants des effets d’échelles [19], qu’ils peuvent fortement interagir entre eux aux différentes échelles [20] et ne fonctionnent pas nécessairement de façon homogène dans
14
l’espace et dans le temps [21]. L’hyper-diversité locale des FTH, qui peuvent abriter plus de 300 espèces d’arbres par hectare dans les cas les plus extrêmes, se traduit aussi par une diversité fonctionnelle très importante, mal connue dont les conséquences en termes de SE ne sont pas clairement évaluées [22]. Décrire la variabilité spatiale des écosystèmes et de leur fonctionnement aux échelles intermédiaires, en comprendre les déterminants et partager ces informations avec les acteurs sont donc autant d’étapes à franchir si l’on souhaite réussir la gestion durable des FTH sans en rester aux invocations ou aux principes généraux [18].
FIGURE 2 : REPRESENTATION DES INTERACTIONS ENTRE LES ECHELLES ECOLOGIQUES ET INSTITUTIONNELLES POUR DIFFERENTS SERVICES ECOSYSTEMIQUES
Inspiré des figures 1 et 2 de [23]. Les échelles opérationnelles (i.e. celles auxquelles s’opèrent la majorité des fonctions et s’effectuent la majorité des actions) sont surlignées.
15
I.1.c
La caractérisation des écosystèmes : une démarche
essentielle pour réussir la gestion durable des FTH
D’importants investissements ont été engagés ces dernières années pour mieux comprendre les processus écologiques en œuvre dans les FTH et leur conséquences en termes de SE à l’échelle globale. L’établissement et l’instrumentation de plusieurs centaines de placettes permanentes de 1 à 50 ha réparties sur toute la ceinture tropicale ont permis de mieux comprendre le fonctionnement des FTH à l’échelle locale. La mise en réseaux de ces placettes (CTFS [24], RAINFOR [25], ATDN [26]) apporte une vision de plus en plus claire sur les tendances globales des FTH à des échelles continentales et leur rôle dans les grands cycles biogéochimiques. Cependant au-delà des processus écologiques élémentaires, la diversité des structures forestières et des compositions des communautés biotiques jouent aussi un grand rôle dans le fonctionnement des écosystèmes et la variabilité spatiale des SE [18]. De fait, quelques sites de référence faisant l'objet d'un suivi fin ne peuvent suffire. Les connaissances scientifiques concernant l’hétérogénéité et le fonctionnement des FTH aux échelles opérationnelles intermédiaires font encore aujourd’hui défaut. C’est pourtant aux niveaux des écorégions, des paysages et des écosystèmes que les principales décisions d’aménagement du territoire qui orientent les stratégies de gestion se prennent et c’est au niveau des paysages, des écosystèmes et des parcelles que les actions de gestion opèrent le plus généralement [27].
Deux facteurs peuvent expliquer les difficultés à acquérir les connaissances scientifiques à ces échelles stratégiques pour la réussite de la mise en œuvre de la gestion durable des FTH : premièrement le coût et la difficulté d’accès au terrain qui limite fortement la collecte des données à ces échelles ; deuxièmement la difficulté à définir les échelles d’étude les plus pertinentes dans cet écosystème complexe où la structuration spatiale et l’interdépendance entre les échelles est particulièrement difficile à appréhender [20].
Le concept d’habitat naturel qui est une des pierres angulaires de la mise en œuvre de la gestion durable des écosystèmes en milieu tempéré peut permettre quant à lui de dépasser la complexité inhérente aux effets d’échelles et de mieux appréhender la variabilité spatiale des FTH , de leur fonctionnement et des conséquences sur les SE [18]. Définit comme « une portion de l’espace au sein de laquelle les conditions de milieu et les ressources satisfont aux besoins de plusieurs espèces» [28], le concept d’habitat naturel s’adapte à l’ensemble des échelles du biome (macro-habitats) à la parcelle (micro-habitats) [29]. Il est centré prioritairement sur l’étude des relations entre les différentes composantes de l’écosystème (assemblage des espèces au sein des communautés, influence de l’environnement sur les populations,…) et définit ainsi un cadre d’étude adaptable aux différentes fonctions étudiées. Intégrant les
16
conditions environnementales mais aussi la structure et la composition des communautés qui s’y reproduisent et s’y maintiennent [30], il permet de caractériser les principaux SE de support dépendant de la biodiversité ou du contexte abiotique [31]. Il permet de développer des indicateurs synthétiques sur l’intégrité des SE aux différentes échelles opérationnelles à travers la définition d’espèce-parapluies, d’habitats clefs, etc... [32].
Pour surseoir aux limitations de moyens et collecter une masse de données suffisante à une telle approche écosystémique, de nouvelles démarches scientifiques sont aujourd’hui engagées. De plus en plus, la communauté scientifique mobilise de nouveaux acteurs de la recherche tels les para-taxonomistes (e.g. [33]) ou les communautés rurales locales détentrices de savoirs traditionnels (e.g. [34,35]). Elle utilise des sources de données alternatives telles les inventaires forestiers (e.g. [36]) ou des méthodes dites de « Rapid Assessment » (e.g. [37]). Ce changement de paradigme ouvre de nouvelles possibilités mais nécessite aussi une adaptation des outils d’analyse à ces nouveaux types de données.
Le développement de la télédétection en général et de son volet satellitaire en particulier, offre aussi de nouvelles facilités pour appréhender la dimension spatiale de la diversité des écosystèmes [38]. En offrant une nouvelle perception de l’environnement sur de larges emprises (i.e. > centaines de km²), elle fournit les moyens d’aborder objectivement les effets d’échelles et la notion d’interaction entre ces échelles.
La caractérisation multi-échelle des habitats naturels s’appuyant sur ces méthodes d’inventaires rapides est à même de fournir des éléments essentiels à la compréhension du fonctionnement des FTH aux échelles intermédiaires. Leur cartographie à travers des modèles prédictifs ou des approches basées sur la télédétection peut apporter une meilleure appréhension de la variation des SE aux échelles opérationnelles. Au-delà de son intérêt pour la gestion des SE, cette approche descriptive des assemblages au sein des communautés et des relations avec les facteurs environnementaux peut permettre d’apporter des éléments de compréhension sur l’écologie forestière et l’importance relative des mécanismes à l’origine de la répartition des espèces. C’est le travail que j’ai mené au cours de cette thèse, à l’échelle de la Guyane française, en m’appuyant sur les données de deux récents projets de recherche essentiellement focalisés sur les arbres forestiers, les plantes des sous-bois, les sols et la grande faune (projet ECOTROP « Paysages et Biodiversité » [39], projet PO-FEDER « Habitats » [40]).
17
I.2
La Guyane française et son cadre régional
I.2.a
Le Bouclier guyanais sous-région amazonienne
Situé sur la côte Nord-Est de l’Amérique du Sud, le Bouclier guyanais constitue un vaste ensemble de 2.3 millions de km², assis sur un craton Précambrien de plus de 1.7 milliard d’années. Il couvre la totalité du Guyana, du Suriname, de la Guyane française ainsi qu’une partie du Venezuela, du Brésil (Etats d’Amapa et du Para) et de la Colombie [41]. La forêt tropicale humide recouvre près de 70% de sa surface et constitue, avec le reste du massif Amazonien auquel il est contigu, le plus grand massif forestier tropical naturel au monde avec encore aujourd’hui plus de 6 millions de km² non artificialisé [5]. Malgré sa position périphérique et sa moindre contribution en termes de surface, le massif des Guyanes présente un très grand intérêt dans ce vaste ensemble du fait de nombreuses particularités en termes de composition floristique et faunistique, de structure forestière et de fonctionnement écosystémique.
TABLEAU 1:LE CONTEXTE FORESTIER GUYANAIS ET AMAZONIEN EN QUELQUES-CHIFFRES
Caractéristiques des forêts Amazonie Plateau des Guyanes
Guyane
française Sources Surface de forêt tropicale
humide (en km²) 6 millions 1,6 million 80 000 [5] Nombre estimé d’espèces
d’arbres (DBH>10cm) 15 970 4 581 >1800 [42] + [43] Diversité locale (α-Fisher sur
1ha et DBH>10cm) # 68 33 84 [44]
Biomasse épigée (en t.ha-1 avec
DBH>10cm) $ 287 387 (393) [45]
# valeurs moyennes calculées à partir du tableau de l’annexe 1 de [44] - $ valeurs moyennes calculées à partir des informations supplémentaires de [45] avec seulement 3 localités pour la Guyane française (valeur entre parenthèses)
Avec un nombre d’espèces d’arbres estimé à près de 4 600 [42], une alpha-diversité modeste par rapport à d’autres régions amazoniennes [44], la région n’est pas considérée comme un hot-spot [7] ni une aire de conservation prioritaire pour sa flore [46,47] malgré un endémisme assez marqué [48]. Cependant elle recèle une grande richesse faunistique, notamment en mammifères [49]. Elle fait partie des 200 écorégions d’intérêt représentative de la biodiversité mondiale [9] du fait de sa composition spécifique très originale à l’échelle amazonienne avec une dominance de Fabaceae, de Lecythidaceae et de nombreuses espèces de fin de succession à bois très denses et graines lourdes [50] indicatrices d’un degré de maturité sylvigénétique avancé. La stature des forêts guyanaises est elle aussi remarquable avec une hauteur de canopée dépassant fréquemment les 45m, bien supérieure à
18
celle des autres forêts amazoniennes [51], et une très forte concentration de biomasse sur pied estimée à plus de 380 t.ha-1 en moyenne [45]. De fait, le Plateau
des Guyanes abrite une grande partie des espèces les plus contributrices au stock de carbone amazonien [52], notamment plusieurs espèces hyper-dominantes dont l’aire de répartition est concentrée sur cette région et sont fréquemment exploitées pour leur bois telles Eperua falcata, Dicorynia guianensis, Qualea rosea, Chlorocardium rodiei ou Vouacapoua americana [42,53].
I.2.b
La diversité forestière de la sous-région guyanaise
Au-delà de ces traits généraux tirés des principales méta-analyses réalisées à l’échelle globale ou amazonienne, la variabilité intrinsèque des forêts guyanaises reste largement méconnue. La flore des Guyanes, est certes l’une des mieux documentées de la région [54] grâce à l’intensification constante des campagnes de prospection depuis plus de deux siècles et grâce à un effort d’identification très élevé [55,56] cependant la distribution des espèces et la variabilité de leur assemblage en communauté est encore peu documentée à l’exception notable du Guyana (GY) qui a été remarquablement bien couvert par des inventaires forestiers dans les années 1960-1970 et intensément étudié d’un point de vue scientifique [57-59]. Les forêts de la moitié sud de la région couvrant l’Amapa (AM) et les ¾ de la Guyane française (GF) et du Suriname (SR), sont cependant moins documentées et restent aujourd’hui décrites sous le terme générique de forêt de la « Pénéplaine du Sud »[60] ou « forêt des terres hautes » [61]. Ce secteur forestier apparemment homogène (Figure 3) présenterait cependant une très forte diversité spécifique comparé aux forêts de la plaine côtière et aux forêts sur sable blancs qui, à l’inverse, présentent une faible diversité locale mais des formations contrastées liées à des phénomènes d’endémisme, de dominance locale (concernant plusieurs espèces de Caesalpinoideae) et de forts filtres édaphiques [60].
L’observation des canopées forestières par télédétection met aussi en évidence une certaine hétérogénéité de la structure forestière au sein de la région [62] avec des forêts plus hautes et plus régulières sur le nord de la Guyane française, l’Amapa et les plateaux centraux du Guyana, contre des forêts aux canopées plus ouvertes et irrégulières sur le Suriname, le Sud de la Guyane française et l’est du Guyana (Figure 4).
19
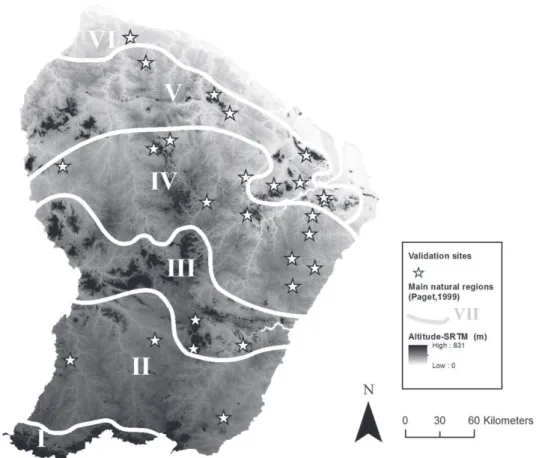
FIGURE 3 : CARTOGRAPHIE DES PRINCIPALES REGIONS FORESTIERES DU BOUCLIER DES GUYANES PROPOSEES PAR TER STEEGE &ZONDERVAN TIREE DE [60]
FIGURE 4 : CARTOGRAPHIE DES TYPES DE PAYSAGES FORESTIERS TIREE DE [62] REALISEE PAR TELEDETECTION
20
I.2.c
Contexte socio-économique en Guyane française
Avec ses 85 000 km² la Guyane est le plus petit territoire de la région Amazonienne, mais son massif forestier représente plus d’un tiers de la surface forestière sous administration française (16,5 millions d’hectares en métropole). Quasi-intégralement domaine privé de l’Etat français, elle représente près des deux tiers des forêts publiques (4,7 millions d’hectares) et est la seule FTH sous la responsabilité d’un état membre de l’Union Européenne.
Pour répondre aux déclarations de l’accord de Rio, la France s’est engagée dès 1992 à mettre en œuvre une gestion durable et multifonctionnelle sur ce territoire d’exception : une mission qu’elle a confiée à l’Office National des Forêts (ONF) et aux services ministériels en charge de l’écologie (DEAL ex DIREN)[63]. Cet engagement s’est concrétisé par trois actions importantes : (i) l’abandon des permis d’exploitation forestière dès 1994 au profit de la mise en place d’aménagements forestiers, première étape d’une démarche aboutissant à la certification de la gestion forestière en novembre 2012 ; (ii) la création le 27 février 2007 du plus grand Parc National français au terme d’une démarche engagée dès 1993 ; (iii) l’adoption le 28 juillet 2005 d’un Code Forestier adapté à la Guyane renforcé par la délimitation d’un Domaine Forestier Permanent (DFP) en 2008. Aujourd’hui le massif forestier guyanais bénéficie d’un statut réglementaire et de vocation stable sur plus de la moitié de sa surface (Figure 5) comprenant 20 300 km² de protection forte et intégrale (zone cœur du Parc Amazonien de Guyane - PAG), 24 000 km² à vocation multiple intégrant une part importante de production (Domaine Forestier Permanent géré par l’ONF), 13 600 km² à vocation multiple devant concilier développement et protection (zone d’adhésion du PAG).
Ce cadre général, n’exclut cependant pas les questionnements concernant l’aménagement du territoire et les conflits d’usage, notamment face aux activités minières, et aux besoins de développement agricole et énergétique. En effet, l’activité aurifère, en plein essor en Guyane depuis une quinzaine d’années comme sur le reste du Bouclier Guyanais [64], n’est pas exclue du DFP ni de la zone de libre adhésion du PAG ce qui amène fréquemment les gestionnaires de ces domaines à devoir défendre ou justifier la vocation forestière de secteurs à potentiel minier. Par ailleurs, les choix d’aménagement du territoire restent ouverts dans la zone de libre adhésion du PAG où le développement de secteurs d’exploitation forestière ou agricole n’est pas exclu. Le DFP de l’ONF doit aussi à terme être intégralement aménagé ce qui implique d’effectuer des choix de vocations prioritaires et des compromis entre les différents SE.
Force est de constater que les connaissances nécessaires à l’optimisation de ces choix sont encore minces, la variabilité du massif forestier guyanais vis-à-vis des
21
différents SE étant encore très mal connue à l’échelle du territoire. Ces lacunes ont été particulièrement criantes lors de la réalisation du Schéma Départemental d’Orientation Minière (SDOM) en 2007 qui a nécessité la proposition de zones prioritaires de protection pour lesquelles peu d’éléments de diagnostic étaient alors disponibles, si ce n’est de simples listes d’espèces et les zonages réglementaires déjà existants [65]. L’acquisition d’informations scientifiques précises sur la diversité des écosystèmes forestiers et de leur fonctionnement est encore à ce jour une étape essentielle pour asseoir une gestion durable raisonnée et efficace sur ce massif.
FIGURE 5 : CARTOGRAPHIE DES ZONES FORESTIERES REGLEMENTAIRES EN VIGUEUR EN GUYANE
En vert le Parc Amazonien de Guyane et en gris le Domaine Forestier Permanent géré par l’Office National des Forêts
22
I.3
Cadre scientifique et structure de la thèse
I.3.a
Objectifs et questions de recherche
Les travaux réalisés dans le cadre de cette thèse visent à étudier la diversité des écosystèmes forestiers de Guyane, les facteurs déterminant leur variabilité et les conséquences de cette variabilité en termes de services écosystémiques. Ils ont pour finalité de fournir aux gestionnaires et aménagistes locaux de nouveaux outils permettant d’appréhender la diversité écologique des forêts. Mon approche se base sur la définition et la caractérisation d’habitats forestiers à partir de télédétection et de l’analyse de données d’inventaires multidisciplinaires rapides recueillies entre 2006 et 2013 dans le cadre du projet de recherche HABITATS piloté par l’Office National des Forêts (ONF) et l’Office Nationale de la Chasse et de la Faune Sauvage (ONCFS). Au cours de ces travaux j’ai poursuivi trois objectifs principaux déclinés en autant de question de recherche.
Le premier objectif vise à combler ce que l’on appelle le « Wallacean shortfall » (d’après Lomolino in [66]), c’est-à-dire le déficit de connaissance en termes de distribution régionale des espèces et de variation de composition des communautés (ci-après désigné par le terme de bêta-diversité dans son acceptation la plus large [67]). La bêta-diversité forestière présente-t-elle des organisations spatiales détectables et à quelles échelles ? Telle sera la question de recherche relative à ce premier volet. Pour répondre à cette question je me suis inscrit dans un cadre statistique de décomposition de bêta-diversité combinant mesure de la diversité et analyses spatiales [68].
Le deuxième objectif vise à rechercher les mécanismes écologiques à l’origine de l’organisation spatiale de la bêta-diversité observée. Il s’agira principalement de faire la part entre l’influence des processus déterministes (ou effets de niches) liés aux filtres environnementaux et les processus de stochasticité démographique liés à la dispersion des espèces (ou effets neutre).
Ces deux premiers objectifs permettront de déterminer à quelles échelles et selon quelles clefs d’entrée il est possible de définir des habitats forestiers représentatifs de cette bêta-diversité.
Enfin le troisième objectif vise à évaluer les conséquences de cette diversité sur les principaux SE pris en compte par les gestionnaires forestiers guyanais, notamment le stockage du carbone (service de régulation) et l’approvisionnement en bois (service de production). « Quelle influence la biodiversité des forêts guyanaises exerce-t-elle sur les fonctions de production de bois et de régulation du carbone? » sera la dernière question de recherche abordée.
23
I.3.b
Cadre méthodologique
I.3.b.1 Partitionner la bêta-diversité dans un cadre statistique robuste
La bêta-diversité spécifique des communautés d’arbres forestiers présente-t-elle des organisations spatiales détectables et à quprésente-t-elles échprésente-t-elles ? Dans quprésente-t-elle mesure des facteurs écologiques facilement cartographiables à grande échelle, peuvent-ils rendre compte de ces variations ? Pour répondre à ces questions je me suis inscrit dans un cadre statistique combinant décomposition de la diversité et analyses spatiales [68].
La diversité peut en effet se décomposer en une composante alpha (ou diversité locale) et une composante bêta (relatif aux changements de composition entre localités) [69]. La différence de nature entre bêta et alpha (la première mesurant une variation et la seconde une quantité) est à l’origine de nombreuses polémiques quant à la façon de définir la bêta-diversité (diversité inter-localité, taux de remplacement le long d’un gradient, etc.) et de modéliser sa relation avec la fraction alpha (décomposition additive ou multiplicative) [67,70]. La profusion des indices permettant de mesurer la diversité (basé sur le nombre d’espèces présentes ou en tenant compte de leur abondance, …) et la nature éminemment incomplète des échantillonnages spécifiques dans des milieux aussi diversifiés que les forêts tropicales humides complexifient encore le problème [71]. Ces questions relatives à la définition et à la mesure de la bêta-diversité sont loin d’être tranchées et font encore l’objet d’une littérature foisonnante.
Parmi les différentes approches proposées pour mesurer la bêta-diversité, j’ai adopté celle basée sur la mesure de variance de tables d’occurrences (i.e. somme des carrées = TSS), qui découlent des tables de relevés classiques (relevés d’abondances d’espèces par localités). Ce cadre proposé par Pélissier et collègues [72] définit la bêta-diversité comme l’inertie inter-relevés de la table (ou part de variance « expliquée » par les relevés = MSS) qui, moyennant différentes pondérations, permet d’obtenir les équivalents de différentes mesures de bêta-diversité. C’est un strict équivalent à toutes les étapes de la décomposition en ce qui concerne Simpson (ou entropie d’ordre 2) et une équivalence formelle au niveau de la table des relevés, pour ce qui est de la Richesse (entropie d’ordre 0) et de Shannon (entropie d’ordre 1). Le système des pondérations entre espèces permet donc de définir plusieurs schémas de décompositions accordant une importance décroissante aux espèces rares selon que la variance totale de la liste des occurrences déplace la référence de l’entropie d’ordre zéro vers l’entropie d’ordre 2 [73].
Au-delà de cette propriété, cette approche possède plusieurs avantages. Premièrement, elle permet une décomposition additive de la diversité (telle que proposée par Lande [74]) et s’adapte ainsi aux tests classiques de comparaison de
24
variance (ANOVA), le ratio bêta-diversité sur diversité totale pouvant être considérée comme une mesure de R² [75]. Deuxièmement, la bêta-diversité peut faire l’objet d’une décomposition multifactorielle, les relevés pouvant être regroupés par facteurs environnementaux ou en fonction de leur relation spatiale à partir de matrices de voisinages ou de classes de distances entre relevés [73]. De plus, au-delà de la seule décomposition de la variance totale de la liste d’occurrences et de la table des relevés, le cadre définit de façon cohérente des familles d’analyses multivariées (i.e. AFC et ANSC) qui permettent la réalisation de typologies duales espèces-relevées, accordant plus ou moins de poids aux espèces rares (AFC ou Analyse Factorielle des Correspondances) ou aux espèces abondantes (ANSC ou Analyse Non-symétrique des Correspondances). Les analyses de covariance, de même nature mais introduisant des tables de variables externes (l’Analyse Canonique des Correspondances ou CCA équivalent de l’AFC et une forme d’Analyse de Redondance RDA appelée ANSC à variable instrumentale) entrent aussi dans ce cadre (table2). TABLEAU 2: CORRESPONDANCE ENTRE LES METHODES D’ORDINATION ET LA DECOMPOSITION DE LA DIVERSITE A PARTIR DES TABLES D’OCCURRENCES TRANSFORMEES
Méthodes d’ordina-tion Mesures de diversité et partition-nement Transformation de la table d’occurence (yai) et pondération des
relevés (δa) et des espèces (wi) à
partir des fréquences (pai)
Indice
correspondant
AFC TSS/n=S-1 yai=pai/ pa+ ; δa=pa+ ; wi=1/ p+i S = Richesse
CCA R²=MSS/TSS
ANSC TSS/n=E yai=pai/ pa+ ; δa=pa+ ; wi=1 E =
Simpson-Gini ANSCIV
(RDA)
R²=MSS/TSS
pai correspond à la fréquence relative de l’espèce i sur le relevé a –p a+ à la somme des
fréquences sur le relevé a et p+i à la somme des fréquences de l’espèce i – TSS est la somme des
variance de chaque espèce (i.e. ΣiwiΣa(yai-y.i)² avec y.i la fréquence moyenne de l’espèce i) et
MSS la variance expliquée par la variable instrumentale (i.e. ΣiwiΣa(ŷai-y.i)² avec ŷai la
fréquence estimée de l’espèce i sur le relevé a en fonction de la variable instrumentale)
Si cette approche peut soulever certains problèmes conceptuels vis-à-vis de certaines propriétés attendues des indices de mesure de la diversité [76] ou de l’indépendance de calcul exigée, par certains, entre alpha et bêta-diversité [70], son utilisation avec des métriques de Simpson est reconnue comme robuste, et cohérente avec d’autres types d’approches, par exemple celle basée sur l’entropie ou, plus directement, avec les fonctions de similarité. Ces dernières sont centrales dans les approches classiques de génétique des populations et sont très facilement connectables avec les paramètres fondamentaux de la théorie neutre (voir plus loin) [77]. Outre que cette approche possède le grand avantage de pouvoir s’insérer dans le cadre des analyses multivariées classiques (analyses de types AFC, ANSC, avec ou sans contraintes – voir ci-dessus), elles peuvent aussi se décliner dans le cadre des
25
analyses spatiales classiques de types variogrammes [78]. La combinaison de ces deux cadres statistiques dégage aujourd’hui de puissants outils de décomposition de la diversité dans un cadre spatialement explicite et connecté à son environnement [68].
I.3.b.2 Expliquer les patrons de diversité par les effets de niche et effets neutres Quelle est la part relative de diversité expliquée par les effets de niches liés aux filtres environnementaux et par les effets neutre ? Cette problématique est récurrente en écologie, notamment des écosystèmes tropicaux dont on cherche à comprendre l’origine de la très forte diversité. Deux théories majeures définissent le cadre conceptuel dans lequel sont abordées les questions relatives aux patrons de diversité (abondances relatives des espèces, diversité phylogénétique, distributions dans l’espace et dans le temps, etc.) : la théorie des niches et la théorie neutre. Elles ont été maintes fois exposées, testées et longtemps opposées mais sont actuellement plus souvent perçues comme complémentaires [79].
Très succinctement, la théorie des niches écologiques, proposée par Hutchinson [80] explique la diversité des communautés par la diversification des « niches », c’est-à-dire des conditions d’accès aux ressources et à l’énergie nécessaire au maintien des populations. Comme aucune espèce ne peut être la meilleure compétitrice dans tous les domaines mais doit se plier à des compromis fonctionnels [81], la survie de leur population va donc dépendre de leur plus ou moins bonne adaptation au milieu, c’est-à-dire la plus ou moins bonne adéquation de leur niche théorique (ou fondamentale) avec l’habitat en place. La niche de chaque espèce serait la résultante de son évolution. Théoriquement, deux espèces occupant la même niche se retrouvent en compétition ce qui doit aboutir inéluctablement à la disparition d’une des deux espèces (exclusion compétitive [82]). De ce point de vue, seule une forte diversification des niches (diversité des traits de vie, variabilité des besoins au cours de l’ontogénie, …mais aussi incluant les interactions biotiques) peut expliquer la coexistence d’un grand nombre d’espèces. La théorie de la perturbation intermédiaire [83], s’inscrit aussi dans ce cadre puisqu’elle prévoit un maximum de diversité dans des conditions de perturbation modérées (en intensité et en fréquence) permettant ainsi le maintien d’une variabilité spatiale et temporelle de l’environnement et des stades successionnels [84].
A l’inverse, la théorie neutre tente d’expliquer la diversification des communautés sans avoir à faire appel une spécialisation des espèces, ni même à toute forme de variance entre individus. Elle démontre que même dans une situation de complète équivalence écologique des espèces, les phénomènes stochastiques de recrutement, dispersion et de mortalité, en présence d’un nombre fini d’individus, sont suffisants pour expliquer une dérive écologique entre communautés. Plus précisément chaque mortalité d’un individu au sein de la communauté à une probabilité (1-m) de laisser
26
place à la naissance d’un nouvel individu d’une espèce déjà présente au sein de la communauté (recrutement). Cependant, il y existe aussi une probabilité m pour que ce nouvel individu provienne d’une espèce extérieure (immigration) provenant de la méta-communauté. Cette probabilité m dépend de la capacité de dispersion des espèces (dispersal limitation). Au sein de la méta-communauté, l’apparition d’une nouvelle espèce (spéciation) participe aussi à cet équilibre avec une probabilité θ. La calibration de ces deux seuls paramètres m et θ suffit à générer des distributions d’abondances d’espèces non-triviales et très souvent non-différentiables des patrons observés en FTH concernant, aux niveaux local et régional, les distributions d’abondances d’espèces, les relations aires-espèces et certaines mesures de béta-diversité [85].
Il n’est guère discutable que des mécanismes relevant du concept général de niche existent dans les écosystèmes considérés et il n’est pas non plus discutable que la dispersion limitée et la stochasticité démographiques façonnent, dans une certaine mesure, la dynamique des populations des différentes espèces [86,87]. La question concernant l’importance relative de ces deux mécanismes potentiels non exclusifs a déjà plusieurs fois été posée en Guyane mais toujours à des échelles locales difficilement généralisables et la question reste donc encore aujourd’hui ouverte. La ligne de force sur laquelle je vais m’appuyer au cours de cette thèse pour aborder cette question est tirée des travaux précurseurs de Paget [88]. Elle suppose une relation forte entre le contexte géomorphologique et la composition forestière, liée à différents mécanismes, probablement interdépendants, et relevant chacun du concept global de niche. La variabilité des formes de reliefs, que l’on peut caractériser à différentes échelles, reflèterait la diversité des conditions environnementales abiotiques (géologique, pédologique, topographique) à diverses échelles et permettrait donc d’expliquer celle des habitats forestiers. Les typologies et cartographies géomorphologiques auraient donc une valeur prédictive sur la distribution des sols et les compositions floristiques [88]. Cette hypothèse a rapidement été adoptée par les forestiers qui ont alors utilisé la géomorphologie comme clef de stratification pour l’analyse écologique des forêts à aménager [89]. La question se pose donc aujourd’hui de la réelle efficacité de cette séduisante approche dont il est impératif de tester formellement la performance en termes de pouvoir explicatif (au sens de la décomposition de la variance floristique des communautés).
I.3.b.3 Analyser les relations entre biodiversité et SE à l’échelle du territoire Quelle influence la biodiversité exerce-t-elle sur les autres SE notamment ceux relatifs à la production et à la régulation ? Les capacités de production et de stockage des écosystèmes sont-elles fonctions de leur diversité ? Peut-on optimiser
27
conjointement les SE de production et la biodiversité ou doit-on rechercher un nécessaire compromis ?
Face à la perte de biodiversité observée dans les écosystèmes perturbés ou transformés par l’homme, ces questions relatives au lien biodiversité-SE s’imposent comme une problématique complexe mais incontournable [90]. De nombreuses expérimentations ont été menées pour tenter de répondre à celles-ci et suggèrent une corrélation positive entre biodiversité et autres SE [91] avec cependant des effets de seuils [92]. L’optimisation des SE nécessiterait donc un certain niveau de diversité, notamment fonctionnelle, permettant de garantir une utilisation maximale des ressources fournies par l’environnement, et une stabilité de la productivité dans le temps [93]. Cependant, la plupart de ces essais ont été menés à des échelles locales, sur des écosystèmes artificialisés et simplifiés à l’extrême (généralement des expérimentations aux champs sur culture agricoles). La généralisation des résultats de ces expérimentations au « monde réel » est donc parfois critiquée voire controversée [94].
Qu’en est-il effectivement pour des écosystèmes naturels et hyper-diversifiés, telle que les écosystèmes forestiers guyanais, et sur des échelles opérationnelles plus larges ? Dans notre contexte la question des relations biodiversité-SE se pose plutôt en termes de stratégie de conservation et de gestion globale des SE à l’échelle du territoire [95] plutôt que d’optimisation des multiples SE à l’échelle locale. La nature des relations entre biodiversité et SE dans les écosystèmes forestiers naturels est aussi à même de nous renseigner sur les conséquences des processus de perturbation ou de secondarisation des forêts sur les SE (e.g. .[96]) mais cet aspect ne sera pas aborder dans cette thèse. L’analyse qui sera menée ne relève pas de la recherche de corrélations à valeur déterministe inhérentes à des mécanismes reliant diversité et SE mais relève d’une analyse de congruence entre services à différentes échelles [97]. Si une corrélation positive entre la biodiversité et les autres SE se confirme dans notre contexte (ce qui est loin d’être évident compte-tenu des effets de saturation observés sur des écosystèmes simplifiés), les stratégies de conservation de la biodiversité peuvent aussi indirectement servir d’autres objectifs comme la séquestration du carbone par exemple (e.g. [98]). La nature de la relation biodiversité-SE peut aussi accentuer ou au contraire détendre les contraintes imposées pour la gestion multifonctionnelle des forêts et des territoires [99]. Cette problématique sera abordée à l’aide d’analyses de covariance (telles que proposées par Kremen [27]) et sera mise en œuvre à l’aide d’analyses multivariées (tel que Grimaldi [100]), permettant d’appréhender les relations entre biodiversité et SE en tenant compte des interactions possibles entre services.
28
I.3.c
Structure du document
Cette thèse est organisée en trois chapitres qui s’appuient essentiellement sur des articles publiés, soumis ou en cours de rédaction. Chaque chapitre est structuré en quatre parties : une introduction, deux parties de développement prenant généralement la forme d’articles et une conclusion spécifique.
Le premier chapitre est intitulé « Diversité et Distribution ». Il débute par une synthèse rapide de l’état des connaissances concernant la diversité des écosystèmes forestiers guyanais et expose les hypothèses dont je me suis inspiré pour organiser ce travail de recherche, notamment celle d’une structuration de la diversité forestière par la forme des reliefs (ci-après désigné sous le terme de géomorphologie). La première partie présente les méthodes que j’ai préalablement développées afin de pouvoir tester explicitement cette hypothèse à partir d’inventaires forestiers rapides. La deuxième partie présente la démarche d’analyse multi-échelle qui m’a permis de tester explicitement les relations entre géomorphologie et composition forestière et de mettre en évidence une forte structuration de la bêta-diversité à l’échelle des paysages.
Le deuxième chapitre est intitulé «Déterminants et Mécanismes ». Il expose les différents mécanismes pouvant expliquer les liens entre forme des reliefs et diversité forestière, et focalise particulièrement sur l’étude des relations entre géomorphologie, sols et végétation. La première partie décrit l’organisation des systèmes-sols en relation avec la forme des reliefs suggérant l’influence d’un effet de filtre édaphique sous-jacent au déterminisme géomorphologique de la diversité forestière. La deuxième partie évalue formellement la contribution de cet effet de filtre édaphique dans le déterminisme de la bêta-diversité et confirme que les effets de niches sont insuffisants pour expliquer à eux seuls la diversité actuelle des forêts guyanaises. Ces résultats sont mis en perspectives avec la bibliographie afin de replacer la diversité observée dans un cadre historique et biogéographique.
Le troisième chapitre est intitulé « Diversité et Services Ecosystémiques». Il vise à appréhender les conséquences sur les SE de la diversité écosystémique précédemment mise en évidence à la lumière des deux précédents chapitres. La première partie, focalise spécifiquement sur l’évaluation et la cartographie du stock de biomasse forestière à partir des inventaires forestiers, comme indicateur du service de régulation du cycle du carbone rendu par la forêt. Dans la deuxième partie, les mesures de biodiversité développés dans le premier chapitre sont mises en perspectives avec cet indicateur biomasse, les stock de carbone du sol évalué dans le deuxième chapitre et de nouveaux indicateurs d’approvisionnement en bois afin de mettre en évidence les compromis entre biodiversité et principaux SE.
29 FIGURE 6 :SCHEMA D’ARTICULATION DE LA THESE.
Les principales données d’entrée sont indiquées dans les rectangles – les différentes étapes d’analyse sont figurées par les cercles – chaque chapitre est indiqué par une couleur différente – les lettres encadrées indiquent les sections ou articles (en annexes) correspondants à l’étape d’analyse – Les italiques précisent les principales méthodes employées
30
Chapitre 2
II
Diversité et distribution
II.1
Introduction au chapitre
II.1.a
Etat des connaissances sur la diversité forestière
guyanaise
L’inventaire de la flore de la Guyane a débuté dès le 18e siècle notamment avec les travaux de Fusée-Aublet, mais c’est essentiellement depuis la création de l’Herbier ORSTOM/IRD de Guyane, par Oldeman dans les années 1980, que les efforts de collecte et d’inventaire se sont accélérés [55]. Aujourd’hui avec environ 4250 espèces d’Angiospermes répertoriées, on estime connaître plus de 85% des plantes à fleurs présentes sur le département. Cependant l'analyse macro-écologique des patrons de distribution d'abondances laisse entrevoir un large déficit de connaissance des espèces d’arbres les plus rares [42]. De plus, si la flore guyanaise est aujourd’hui globalement parmi les mieux connues, une large partie du territoire est encore sous-échantillonnée [65]. La distribution des espèces, leurs préférences stationnelles, leurs modalités d’assemblage en communauté restent encore largement méconnues – c’est ce que l’on appelle le déficit « Wallacéen »[66].
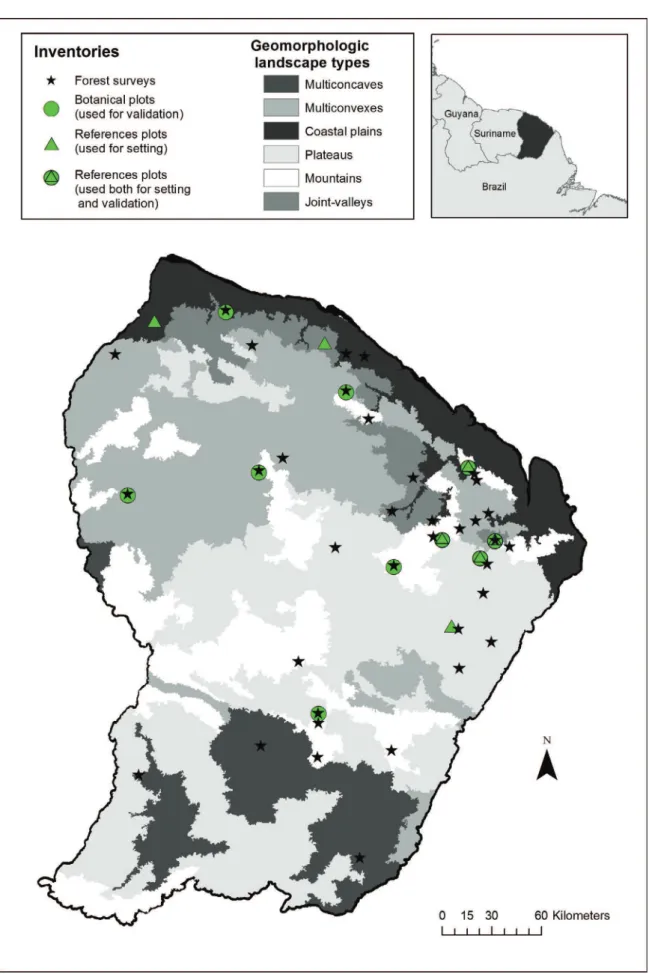
Les espèces d’arbres représentent à elles seules près de 40% de ce cortège floristique forestier (1600 espèces recensées en Guyane [43]) et concentrent l’essentiel de la biomasse forestière [101]. De nombreuses études ont déjà été menées depuis les années 1970 pour tenter de comprendre comment s’assemblent ces nombreuses espèces au sein des communautés hyper-diversifiées et comment ces assemblages réagissent à leur environnement. Cependant l’essentiel de ces études se sont jusqu’à présent focalisées sur l’échelle locale au sein de dispositifs expérimentaux très instrumentalisés, dont la valeur représentative est encore mal cernée compte-tenu de leur concentration dans la partie nord du territoire (Figure.7 – Paracou, Piste de St Elie, Nouragues).
II.1.a.1 Un effet de niche marqué à l’échelle locale
La diversité de la forêt guyanaise a souvent été abordée sous le prisme des relations sol-végétation à l’échelle locale (i.e. celle de placettes de quelques hectares) [28,88,102-106]. Ces études ont mis en évidence l’influence significative des effets de niche (c’est-à-dire l’influence des conditions environnementales sur la capacité de survie des espèces via la modification des conditions d’accès aux ressources [80]). L’influence de la qualité du drainage interne des sols sur l’abondance des
31
espèces a été plusieurs fois démontrée sur les différents dispositifs guyanais avec cependant des résultats variables voire parfois contradictoires en terme d’autécologie des espèces (voir tableau 3). Le contraste entre le cortège floristique des bas-fonds sur sols hydromorphes et celui des forêts de terre ferme est relativement bien marqué et relativement stable entre les sites, avec quelques espèces indicatrices évidentes. Les fortes contraintes édaphiques induites par l’engorgement permanent des sols entraînent non seulement une sélection des espèces les mieux adaptées à l’anoxie mais favorisent aussi le cortège des espèces pionnières et/ou héliophiles du fait d’une dynamique de perturbation autoentretenue [28,107] : l’engorgement du sol limite l’enracinement ce qui maintient une ouverture permanente de la canopée du fait de la fréquence des chablis ; le fort éclairement favorise les espèces pionnières héliophiles à courte durée de vie ce qui entretient par ailleurs la mortalité à un taux élevé ; la chute des arbres depuis les pentes dominantes vers les bas-fonds en aval participent aussi à cette dynamique.
FIGURE 7 : CARTOGRAPHIE DES SITES D’ETUDES DE LA DIVERSITE FORESTIERE MIS EN PLACE EN