Prior selfing and the selfing syndrome in animals: an experimental approach in the freshwater snail Biomphalaria pfeifferi
12
0
0
Texte intégral
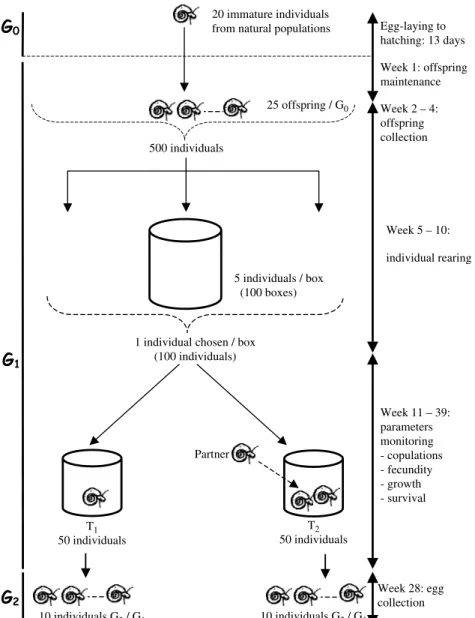
Figure
Documents relatifs