Interplay of DNA supercoiling and catenation during the segregation of sister duplexes
Texte intégral
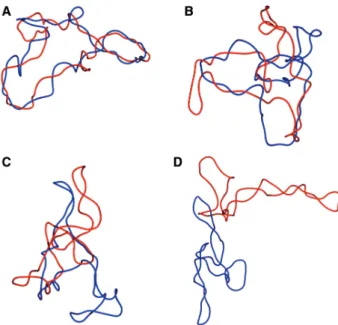
Figure






Documents relatifs
While the Upper Danube is moderately polluted, the tributaries in the Middle and Lower Danube are heavily polluted; however, dilution keeps the Danube critically
tive chromatin fraction from round spermatids after 1.5 rain of digestion, lane C same as lane B but after 3-min digestion, lane D MNase-sensitive
Alors que la vie se poursuit, que les étoiles brillent dans la nuit, la lune quant a elle semble disparue.. Tel une lueur qui se cache, de peur de révéler
Alors qu'elle suit tranquillement sa voie Elle n'a aucune idée sur tes peine etjoie Elle se souvient de toi par erreurs Ta présence éveil sa fureur. Tu vis une vie
Being based in an Alpine country, Botanica Helvetica has traditionally included a high proportion of studies from the Alps or other mountain regions.. The fraction of papers
Not only vascular grafts but also septal defect-occlusion devices interact with blood components (complement, thrombocytes, granulocytes) and serve as an attractive site for
[r]
