Characterisation of the diversity of Fusarium species and their mycotoxigenic potential on French cereals
Texte intégral
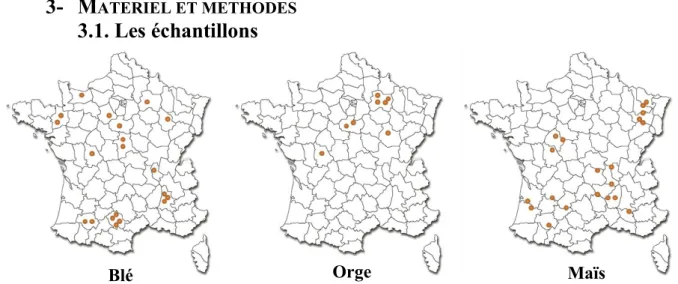
Figure









Outline
Documents relatifs
Les gènes NDM et OXA-48 (famille) ont été retrouvés simultanément chez neuf souches soit six souches d’E.. coli et trois souches
Il est établi qu'il existe une corrélation relativement stricte entre l'espèce à laquelle appartient une souche de champignon et les mycotoxines que cette souche peut produire
Metal Tolerance Protein 1 (MTP1) based on its role in Zn accumulation is considered to be a potential candidate to be expressed in cereals for their Zn biofortification..
[r]
From this study, the large influence of the envelope properties on the flexibility potential was highlighted: the storage efficiency for upward modulations and the rebound rates
L’étude de l’utilisation énergétique de 4 Céréales, effectuée sur 86 agneaux en croissance par estimation de la composition corporelle, indique que la valeur
Influence de la forme de présentation (ensi- lage, déshydratation) d’une céréale immature sur l’efficacité métabolique de l’urée chez le taurillon. en
basse température, si elle est aisée pour l’orge, l’est moins pour le maïs en raison des différences importantes dans les vitesses de dessiccation et les taux
