HAL Id: hal-02811269
https://hal.inrae.fr/hal-02811269
Submitted on 6 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Influence de la densité des tiges et de la proportion du
mélange hêtre/érable sur les caractéristiques
fonctionnelles des feuilles et la croissance de ces deux
espèces
Lucile Demaret
To cite this version:
Lucile Demaret. Influence de la densité des tiges et de la proportion du mélange hêtre/érable sur les caractéristiques fonctionnelles des feuilles et la croissance de ces deux espèces. Sciences du Vivant [q-bio]. 2011. �hal-02811269�
2010-2011
MASTER FAGE
Biologie et Ecologie pour la Forêt, l’Agronomie
et l’Environnement
Spécialité
Fonctionnement et Gestion des Ecosystèmes
Influence de la densité des tiges et de la proportion
du mélange Hêtre/Erable sur les caractéristiques
fonctionnelles des feuilles et la croissance de ces
deux espèces
Lucile DEMARET
Mémoire de stage, soutenu à Nancy le 07/07/2011
Maître de Stage : Damien BONAL, Directeur de Recherche
Laboratoire d’Ecologie et d’Ecophysiologie Forestières – UMR 1137
Equipe Fonctionnement intégré de l’arbre et de l’écosystème
INRA – Centre de Recherches de Nancy
54280 CHAMPENOUX
REMERCIEMENTS
Je tiens tout particulièrement à remercier mon maître de stage, Damien Bonal, sans qui ce stage n’aurait pas pu avoir lieu. Merci pour sa disponibilité, son aide précieuse et sa gentillesse.
Merci à Pierre Montpied, pour ses conseils de statisticien, son aide pour la mise en place du protocole et l’échantillonnage sur le terrain.
Merci également à Anthony, pour avoir donné un peu de son temps pour nous aider sur le terrain. Merci aux personnes qui nous ont prêté le matériel.
Finalement, je tiens à remercier le laboratoire d’Ecophysiologie et Ecologie Forestière, au personnel de la cantine et à l’INRA pour son accueil chaleureux.
SOMMAIRE
I. Introduction 1
II. Matériel et Méthodes 2
1) Zone d’étude 2
2) Matériel végétal 3
3) Echantillonnage 3
4) Liste des mesures à effectuer 4
a. Au niveau de l’arbre 5
b. Au niveau de la feuille 5
5) Analyses statistiques des données 6
III. Résultats 6
1) Hauteur 7
2) Circonférence 8
3) Taux de croissance relative en hauteur 8
4) Taux de croissance relative en surface terrière 9
5) Epaisseur des feuilles 9
6) Masse surfacique (LMA) 10
7) Indice de teneur en chlorophylle (SPAD) 10
8) Densité stomatique 11
IV. Discussion 11
1) Croissance et caractéristiques foliaires chez l’érable sycomore 11
2) Croissance et caractéristiques foliaires chez le hêtre 13
V. Conclusion 15
Bibliographie 15
Annexes 17
1
I.
INTRODUCTION
Un peuplement est caractérisé de « mélangé à n espèces » à partir du moment où : « sur le cercle de 25 m de diamètre sur lequel est effectuée la description du peuplement, n espèces ont un taux de couvert libre1 supérieur à 75 % » (Morneau et al., 2008). A l’heure actuelle, deux tiers de la surface forestière française sont constitués de peuplements mélangés, dont 68% en mélange de feuillus. Ces peuplements se répartissent du Nord-Est aux Pyrénées en passant par le Jura et les Alpes (Landmann et al. 2008). Les forêts mélangées ont depuis longtemps attiré l’attention des gestionnaires forestiers, mais aujourd’hui encore, de nombreux questionnements demeurent sur l’intérêt de ces forêts (Landmann et al. 2008) en ce qui concerne le type de gestion, leur résilience face aux changements climatiques, la biodiversité, et leur avantage économique potentiel par rapport aux peuplements purs.
En réponse au contexte économique et environnemental actuel et face aux nouveaux enjeux (Grenelle de l’environnement, protection de la biodiversité, changements climatiques), les modes et les objectifs de la gestion forestière évoluent et s’étendent vers d’autres horizons. Les fonctions non productrices : loisirs, culture, valeur paysagère, biodiversité, sont de plus en plus demandées. Parallèlement à cela, la production de bois reste une fonction majeure des peuplements forestiers. Les gestionnaires se trouvent donc obligés de revoir leurs priorités et leur manière de gérer la forêt, en axant leurs objectifs vers les nouvelles demandes.
Malgré le peu d’études précises menées jusqu’à présent sur ces peuplements, certains avantages non négligeables leur sont conférés (Sardin et al. 2008). Au niveau économique : diversité des produits du fait de la présence de différentes essences ; Au niveau social : peuplements mélangés plus diversifiés donc moins monotones ; Au niveau écologique : diversité des espèces plus importante, contribuant au maintien de la biodiversité et à une meilleure résistance face aux perturbations naturelles. Cependant, les mécanismes et les processus en jeu pour expliciter ces avantages potentiels restent à l’heure actuelle encore très peu connus. Depuis 10-15 ans, les études visant à mieux comprendre ces mécanismes (distribution de ces peuplements, croissance, compétition entre espèces, facilitation, …) et les réponses de ces forêts aux facteurs biotiques ou abiotiques se multiplient (Landmann et al. 2008).
C’est dans ce cadre qu’en 1998, le Laboratoire d’Etudes des Ressources Forêt Bois (LERFOB), en collaboration avec l’ONF, a mis en place une parcelle expérimentale en forêt de Haye avec un mélange Hêtre/Erable associé à un mélange de densité des tiges (d’où le nom de « double clinal » donné à cette plantation). L’objectif principal de ce double clinal était d’étudier l’influence de la densité des tiges et de la proportion de mélange d’espèces sur la croissance en
1
Surface totale des houppiers des tiges de cette essence ayant accès à la lumière sur la surface totale des houppiers des tiges de la strate forestière.
2 hauteur et diamètre des arbres, l’architecture des houppiers, et le phénomène d’auto-élagage des branches. Suite à des campagnes de mesures de 1998 à 2006, des premiers résultats ont été présentés en Juin 2007 (Vinckler et al., 2007). Les premiers résultats n’ont montré aucune influence du mélange et de la densité sur la hauteur des arbres, un effet de la densité des tiges sur le diamètre, et un effet du mélange sur le diamètre uniquement pour le Hêtre. Quatre années après ces premiers résultats, au sein de l’UMR « Ecologie et Ecophysiologie Forestière » dans laquelle j’ai réalisé mon stage, nous nous sommes intéressés à ces mêmes effets, dans une situation où la compétition pour la lumière et les ressources hydriques et en éléments minéraux est maintenant plus forte, ainsi qu’à l’influence de la densité des tiges et du mélange d’espèces sur les caractéristiques fonctionnelles foliaires des arbres.
Les questions abordées sont les suivantes. Les différentes densités de tiges et le mélange d’espèces ont-ils des effets sur l’épaisseur des feuilles, leur densité stomatique, leur masse surfacique, et sur la hauteur, la circonférence, le taux de croissance relatif en hauteur et en surface terrière sur la période 2009-2011 ? Nous avons fait l’hypothèse que la compétition entre les espèces, ainsi que les différences de densité des tiges, pourraient engendrer une disponibilité des ressources différentes, qui se traduiraient par des différences de caractéristiques foliaires en lien avec l’acquisition et l’utilisation de l’eau et des éléments minéraux.
II.
MATERIEL ET METHODE
Afin de répondre aux différents objectifs de ce stage, j’ai tout d’abord consacré plusieurs journées à de la recherche bibliographique afin de m’imprégner du sujet, puis de fin mai à mi juin, nous avons effectué les mesures de terrain et de laboratoire, et enfin les deux dernières semaines de mon stage ont été occupées par l’analyse des données et la rédaction du rapport.
1) Zone d’étude :
La zone d’expérimentation choisie a été plantée en 1998 par l’ONF en forêt domaniale de Haye (54). Le suivi est effectué depuis en collaboration avec l’Unité LERBOB de Champenoux. Sa localisation sur plateaux calcaires procure au peuplement une fertilité de sol élevée (bonne réserve en nutriments et en eau, grande profondeur...), favorable à la croissance des arbres.
Afin de répondre aux questionnements scientifiques posés initialement par les équipes de l’ONF et du LERFOB, la plantation a été réalisée selon deux gradients :
- Un gradient de densité de tiges : de 1000 à 25000 tiges/ha (selon l’axe des abscisses) - Un gradient de mélange Hêtre (Fagus sylvatica) / Erable (Acer pseudoplatanus) : de
3 Ce site représente une superficie de 2600 m², à laquelle une zone tampon (largeur = 10 m) a été ajoutée en périphérie afin d’éviter des effets de bord potentiels qui engendrent des biais sur les mesures effectuées. Il en résulte donc de cette plantation une forêt mélangée équienne au sein de laquelle, chaque individu est numéroté et localisé grâce à des coordonnées géographiques propres.
2) Matériel Végétal
Plusieurs raisons sont à l’origine du choix de ces deux espèces : Hêtre et Erable, pour la plantation. Tout d’abord, leur part du volume en mélange est importante : 82 % des érables sont en mélange et 48% en ce qui concerne le hêtre (Morneau et al., 2008). De plus, le hêtre est l’espèce qui structure les peuplements mélangés dans l’Est de la France. En effet, 15% des peuplements forestiers de Lorraine sont en mélange à base de hêtre, et le plus souvent associé à l’érable sycomore (Bock et Richter, 2005, Decocq et al., 2005). La deuxième raison pour laquelle ces espèces ont été choisies est le fait qu’elles aient une écologie similaire, notamment au niveau de leur affinité pour la lumière et leur vitesse de croissance.
3) Echantillonnage :
Nous avons fait le choix de comparer les situations « extrêmes » suivant le gradient de densité et de mélange, et donc de stratifier notre échantillonnage suivant ces deux gradients. Un échantillonnage aléatoire de la zone dans sa globalité n’aurait pas été représentatif, car il n’aurait pas rendu compte du gradient de densité et du gradient de mélange d’espèces
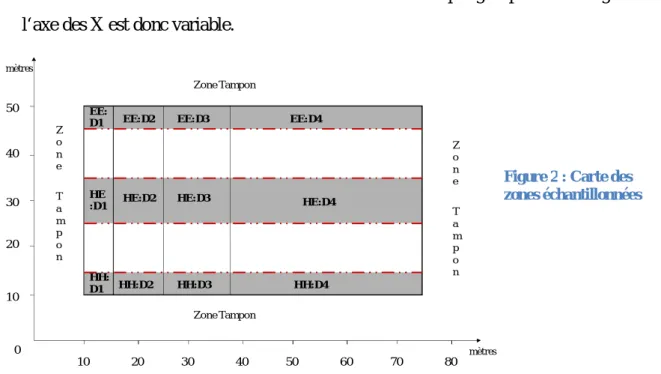
Afin de déterminer les arbres à échantillonner, la zone d’étude a donc été divisée « virtuellement » en 12 sous-zones au sein desquelles on distingue :
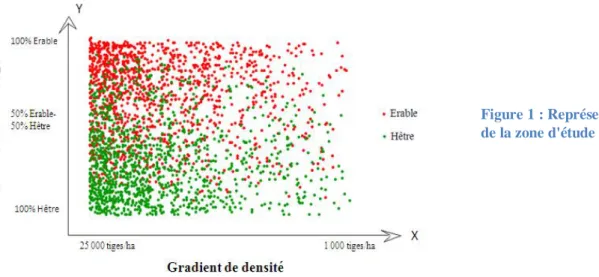
Figure 1 : Représentation de la zone d'étude
4 - 3 groupes de mélange : 100% Erable – 50% Erable/Hêtre – 100% Hêtre. Nous avons fait le choix de travailler sur une bande 5 m de large suivant l’axe des Y pour les groupes de monoculture, et sur une bande de 10 m de large pour le groupe de compétition interspécifique. - 4 groupes de densité de tiges : Ces groupes de densité ont été définis de telle façon à conserver le même nombre d’arbres à l’intérieur de chaque groupe. Leur longueur suivant l‘axe des X est donc variable.
Afin d’avoir un échantillonnage potentiellement représentatif de la variabilité naturelle, 10 arbres ont été échantillonnés par sous-zone en monoculture (hêtre ou érable) et 10 arbres par espèce ont été échantillonnés par sous-zone présentant un mélange des espèces à 50%. Ainsi, au total, nous avons échantillonné 4 groupes de densité * 2 monocultures * 10 arbres, et 4 groupes de densité * 1 composition en mélange * 2 espèces * 10 arbres, c'est-à-dire 160 arbres au total.
Au niveau des sous-zones définies préalablement, le choix des individus à échantillonner a été fait aléatoirement, par tirage au sort grâce au logiciel R (http://www.r-project.org). Une fois cette sélection faite, les individus sélectionnés ont été identifiés à la fois par leur numéro individuel et par leurs coordonnées géographiques.
4) Liste des mesures effectuées
Sur chaque arbre sélectionné, des mesures ont été réalisées au niveau de l’arbre entier et au niveau des feuilles. Les mesures ont été réalisées début juin, quand les feuilles de ces deux espèces étaient totalement formées et matures. Les mesures des caractéristiques de l’arbre ont été effectuées sur le terrain. Les mesures des traits foliaires ont été réalisées sur des feuilles récoltées la journée même ou la veille (afin d’éviter leur desséchement) en laboratoire de l’INRA.
0 10 20 30 40 50 60 70 10 20 30 40 50 80 EE:
D1 EE:D2 EE:D3 EE:D4
HE
:D1 HE:D2 HE:D3 HE:D4
HH:D2 HH: D1 HH:D3 HH:D4 Zone Tampon Z o n e T a m p o n Zone Tampon mètres mètres Z o n e T a m p o n
Figure 2 : Carte des zones échantillonnées
5 a. Mesures au niveau de l’arbre :
- Hauteur : La hauteur de l’arbre (H, m) est définie par la hauteur du dernier bourgeon
terminal. Elle est mesurée avec une canne munie d’une graduation (précision 1 cm).
- Circonférence : La mesure de circonférence (C, cm) du tronc s’effectue à 1.30 m de haut,
dite « hauteur de poitrine », à l’aide d’un ruban gradué.
A partir de ces deux mesures, les taux de croissance relatif en hauteur et en surface terrière ont été calculés. Ces variables permettent de traduire la croissance des individus en prenant en compte leur hauteur et circonférence lors de la campagne précédente (2009).
b. Mesures au niveau de la feuille :
Afin de réaliser ces mesures, les feuilles ont été prélevées sur les arbres, par découpage de quelques branches au sommet de l’arbre, grâce à un échenilloir. Ces prélèvements ont été réalisés sur l’une des branches terminales de l’arbre soumis à un plein éclairement, orientées vers le Sud. 6 à 8 feuilles par arbre ont été prélevées pour les érables, contre 10-12 feuilles pour les hêtres.
- Leaf Mass Per Unit Area (ou masse surfacique) (LMA, g m-2)
La masse surfacique correspond au rapport de la masse sèche de la feuille sur sa surface fraîche. La surface foliaire totale des feuilles est calculée grâce à un planimètre (Modèle Delta-T). Après mesure de la surface foliaire fraîche, les feuilles ont été mises à l’étuve pendant 3 jours à 50° C. Les pesées ont été ensuite réalisées à l’aide de balances avec une précision au mg. Afin d’éviter tout biais de mesure le pétiole des feuilles a été supprimé.
- Epaisseur des feuilles
Cette mesure a été effectuée sur des feuilles fraîches. La mesure est prise au niveau du limbe de la feuille, en prenant soin d’éviter les nervures primaires et secondaires. Elle a été réalisée à l’aide d’un pied à coulisse. Trois mesures par limbe ont été effectuées pour trois feuilles, et la moyenne de ces mesures est ensuite calculée et conservée pour l’arbre.
- Densité stomatique (DS, nombre mm-2)
Cette mesure a été effectuée sur des feuilles fraîches. Trois échantillons ont été prélevés sur trois feuilles différentes pour chaque arbre. La densité stomatique est déterminée par comptabilisation du nombre de stomates dans une zone choisie de la feuille. Le comptage s’effectue sous microscope (grossissement x 400). Afin de réaliser ce comptage, il est indispensable au préalable de réaliser une empreinte de la feuille en disposant une couche de vernis sur la zone à observer, afin de pouvoir voir par transparence l’empreinte des stomates sous le microscope éclairé. Il faut savoir que chez les espèces d’arbres feuillues des régions tempérées, à l’exception du peuplier (LeThiec, comm. perso.),
6 les stomates se trouvent sur la face inférieure de la feuille (feuilles hypostomatiques). Plusieurs répétitions ont été effectuées par échantillon, en prenant soin d’éviter les nervures, et une moyenne des comptages a été réalisée pour obtenir une valeur moyenne de densité stomatique par arbre.
- Indice de la teneur en chlorophylle (SPAD)
Cette mesure s’effectue à l’aide d’un appareil appelé SPAD « Soil Plant Analysis Development ». Cet appareil permet d’estimer un indice de teneur en chlorophylle de la feuille. Trois mesures ont été effectuée par feuille, afin d’obtenir un indice moyen de la teneur en chlorophylle pour chaque arbre.
5) Analyse statistique des données
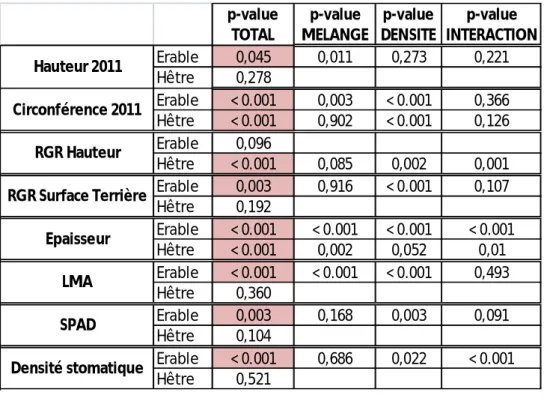
Les données obtenues ont été traitées par une ANOVA multi-facteurs (effet densité, effet compétition, interaction) afin de tester ces effets sur les différentes variables mesurées. Les résultats des « p-value » obtenues sont visibles dans le Tab. 1. Le seuil de 5% a été retenu pour rejeter ou non l’hypothèse nulle. De plus, nous avons comparé les moyennes deux à deux de chaque variable à l’aide d’un test sur les rangs multiples de Duncan. Ce test permet en particulier de regrouper les moyennes en affectant des lettres : les moyennes ayant la même lettre ne sont pas significativement différentes. L’ensemble des analyses a été effectué à l’aide du logiciel de statistiques SAS, en utilisant la procédure « GLM » (SAS-Institue, NC, USA). Un aperçu des résultats sortis de ces tests sous SAS est visible en Annexe.
6) Représentation graphique des données
Les données acquises ont été rentrées dans un classeur Excel. A l’aide de l’outil « Tableau croisé dynamique », les moyennes et erreurs standards ont été obtenues et les graphiques visibles en partie résultats ont pu être établis. Les lettres présentes en étiquettes des barres sont le résultat du test de Duncan sur les rangs multiples.
III.
RÉSULTATS
Par soucis de temps et pour me concentrer sur l’effet du mélange et de la densité de tiges, les caractéristiques des deux espèces ne seront pas comparées. L’astérisque que peuvent comporter certains graphiques signifie que le test statistique du modèle global testé est non significatif pour la variable étudiée.
7
1) Hauteur
Figure 3 : Moyennes des hauteurs, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
Concernant les érables en monoculture, quelque soit la densité de tiges, la hauteur des arbres n’est pas significativement différente (Fig. 3). Par contre, en compétition interspécifique, les érables sont en général plus hauts lorsqu’ils sont en mélange que lorsqu’ils sont en conditions de monoculture (Fig.3). Cette tendance est accentuée dans les faibles densités, où l’écart entre les hauteurs moyennes pour des arbres en monoculture et en compétition est plus important (Fig.3).
Concernant les hêtres, il n’y a pas d’effet apparent de la densité ou du mélange sur la hauteur des arbres (Tableau 1). p-value TOTAL p-value MELANGE p-value DENSITE p-value INTERACTION Erable 0,045 0,011 0,273 0,221 Hêtre 0,278 Erable < 0.001 0,003 < 0.001 0,366 Hêtre < 0.001 0,902 < 0.001 0,126 Erable 0,096 Hêtre < 0.001 0,085 0,002 0,001 Erable 0,003 0,916 < 0.001 0,107 Hêtre 0,192 Erable < 0.001 < 0.001 < 0.001 < 0.001 Hêtre < 0.001 0,002 0,052 0,01 Erable < 0.001 < 0.001 < 0.001 0,493 Hêtre 0,360 Erable 0,003 0,168 0,003 0,091 Hêtre 0,104 Erable < 0.001 0,686 0,022 < 0.001 Hêtre 0,521 Densité stomatique Hauteur 2011 Circonférence 2011 RGR Hauteur RGR Surface Terrière Epaisseur LMA SPAD
Tab. 1 : Résultats des "p-value" de l'analyse de variance multifactorielle pour les différentes variables étudiées BC BC BCBAC C BA BC A 0 100 200 300 400 500 600 700 800 900 D1 Forte Densité D2 D3 D4 Faible Densité H au te u r (e n c m ) ERABLE 0 100 200 300 400 500 600 700 800 900 D1 Forte Densité D2 D3 D4 Faible Densité HETRE
*
HAUTEUR 20118
2) Circonférence
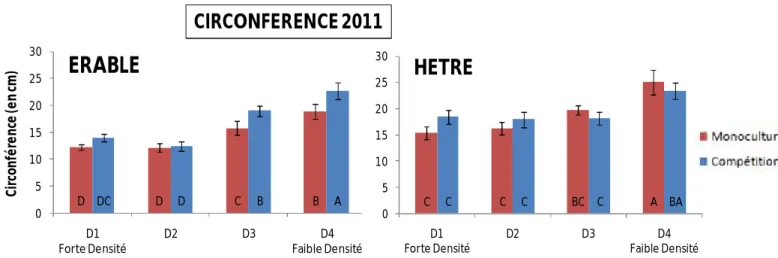
En conditions de monoculture, pour les deux espèces, la densité influence la circonférence des arbres (Tab.1, Fig. 4) : la circonférence est plus importante dans les faibles que dans les fortes densités. Concernant les arbres en compétition, la circonférence des deux espèces est impactée par la densité (Fig.4). Néanmoins, pour le hêtre, seule la classe HE : D4 diffère des autres (Fig.4). Pour l’érable, la circonférence diminue fortement quand la densité augmente (Fig.4).
3) Taux de croissance relatif en hauteur
Concernant les érables, le test statistique est non significatif : il n’y a donc pas d’influence de la densité de tiges et/ou du mélange sur ce paramètre (Tab.1). Pour les hêtres, nous observons un effet significatif de la densité et une interaction densité / mélange (Tab.1). En fait, seule la classe HE : D4, pour laquelle le taux de croissance est supérieur, se distingue des autres (Fig.5).
0 0,05 0,1 0,15 0,2 0,25 0,3 0,35 D1 Forte Densité D2 D3 D4 Faible Densité R G R H au te u r ERABLE
*
B B B B B B B A 0 0,05 0,1 0,15 0,2 0,25 0,3 0,35 D1 Forte Densité D2 D3 D4 Faible Densité HETRERGR Hauteur
D DC D D C B B A 0 5 10 15 20 25 30 D1 Forte Densité D2 D3 D4 Faible Densité C ir co n fé re n ce (e n c m )ERABLE
C C C C BC C A BA 0 5 10 15 20 25 30 D1 Forte Densité D2 D3 D4 Faible DensitéHETRE
CIRCONFERENCE 2011
Figure 4: Moyennes des circonférences, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
Figure 5 : Moyennes du taux de croissance relatif en hauteur, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
9
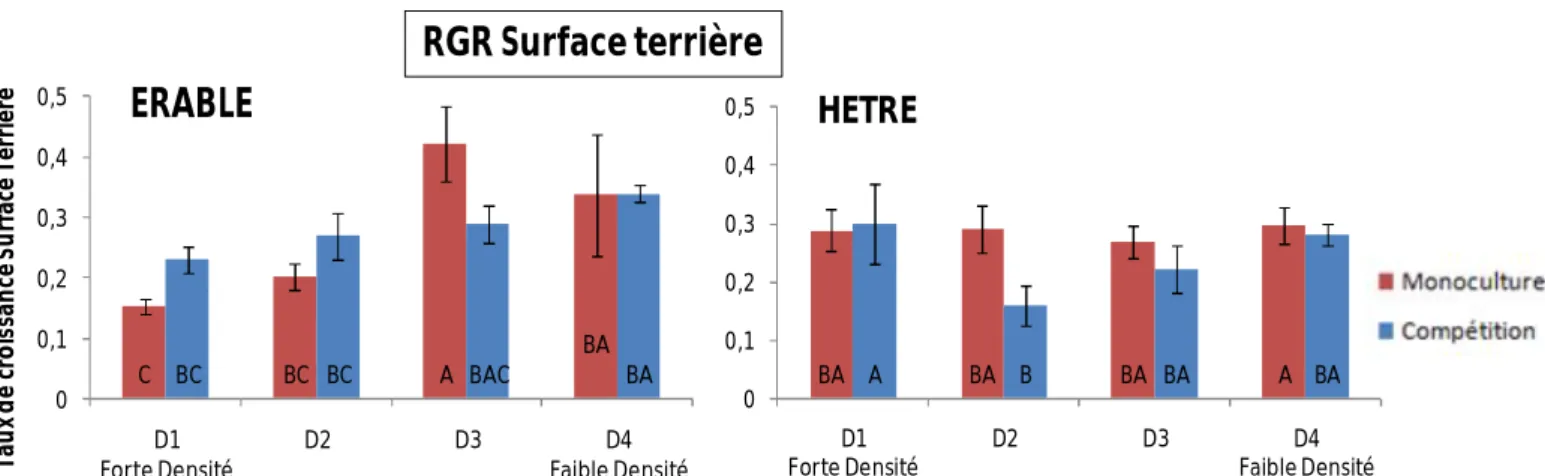
4) Taux de croissance relatif en Surface Terrière
Pour les érables, seul l’effet de la densité est significatif (Tab.1) : les érables en conditions de monoculture montrent un plus fort taux de croissance en surface terrière dans les faibles densités de tiges par rapport aux fortes densités (Fig.6). Concernant les hêtres, aucun effet de la densité et du mélange n’apparaît (Tab.1).
5) Epaisseur des feuilles
Pour les deux espèces, nous observons un effet significatif du mélange, de la densité et une interaction (Tab.1). Concernant les arbres en monoculture, quelque soit l’espèce, il n’y a pas de différence significative entre l’épaisseur des feuilles des différentes classes de densité (Fig.7). Par contre, pour les érables en compétition, l’épaisseur des feuilles diminue quand la densité augmente (Fig.7). Pour les hêtres, seule la classe HE : D4 est différente des autres et montre une épaisseur supérieure (Fig.7). C BC A BA BC BC BAC BA 0 0,1 0,2 0,3 0,4 0,5 D1 Forte Densité D2 D3 D4 Faible Densité Tau x d e cr o is san ce S u rf ac e Te rr iè re ERABLE BA A BA B BA BA A BA 0 0,1 0,2 0,3 0,4 0,5 D1 Forte Densité D2 D3 D4 Faible Densité HETRE
RGR Surface terrière
A B A B A A A A 0,00 0,05 0,10 0,15 0,20 0,25 D1 Forte Densité D2 D3 D4 Faible Densité Ep ai ss e ur de s fe ui ll e s (e n m m ) ERABLE CB CB CB CB C B CB A 0,00 0,05 0,10 0,15 0,20 D1 Forte Densité D2 D3 D4 Faible Densité HETREEPAISSEUR des FEUILLES
Figure 6 : Moyennes des taux de croissance relatif en surface terrière, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
10
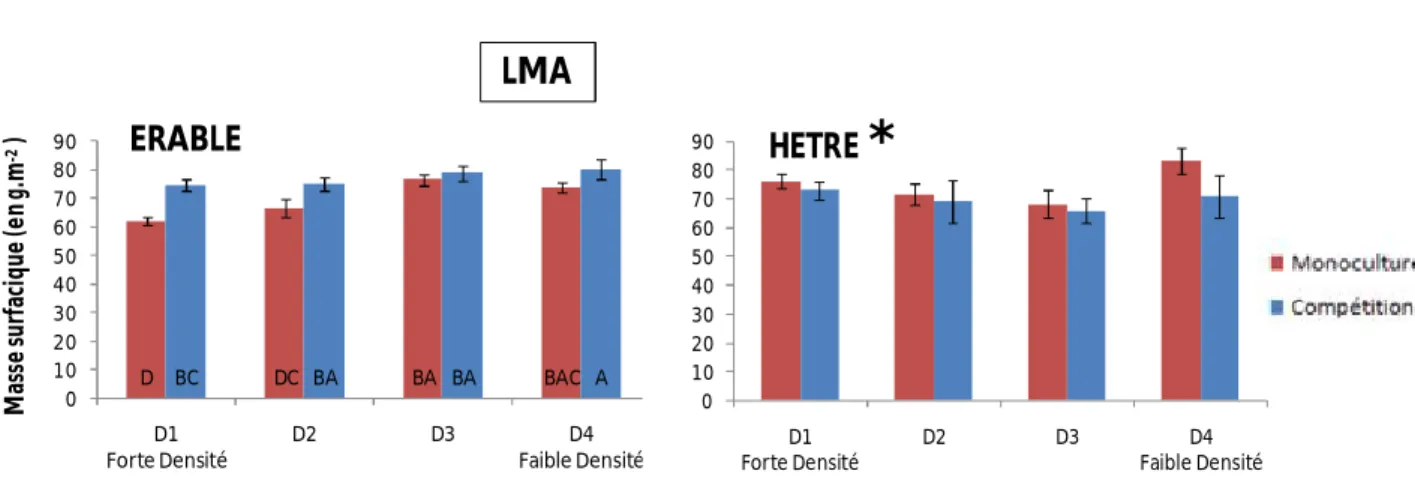
6) Masse surfacique (LMA)
Pour l’érable, nous observons un effet mélange et densité mais pas d’interactions (Tab.1). En monoculture, on peut remarquer une augmentation de la masse surfacique dans les faibles densités (Fig.8). Par contre, pour les érables en compétition, il n’y a une faible différence de LMA entre les différentes classes de densité de tiges (Fig.8). Dans les fortes densités de tiges (D1-D2), on peut observer un LMA plus important pour les arbres en compétition par rapport à la monoculture (Fig.8). Concernant les hêtres, la masse surfacique ne diffère pas significativement selon les densités de tiges ou le mélange (Tab.1).
7) Indice de teneur en chlorophylle : SPAD
Concernant l’indice de teneur en chlorophylle (SPAD) des feuilles, nous n’observons pas de différences entre les cas de conditions de compétition ou de monoculture, quelque soit l’espèce, et un effet significatif de la densité seulement pour l’érable (Tab.1). Néanmoins, pour l’érable, les
D BC DC BA BA BA BAC A 0 10 20 30 40 50 60 70 80 90 D1 Forte Densité D2 D3 D4 Faible Densité M as se s u rf ac iq u e (e n g .m -2) ERABLE 0 10 20 30 40 50 60 70 80 90 D1 Forte Densité D2 D3 D4 Faible Densité HETRE
*
LMA B A A A A A A A 0 5 10 15 20 25 30 35 40 D1 Forte Densité D2 D3 D4 Faible Densité In d ic e d u ta u x d e c h lo ro p h yl le ERABLE 0 5 10 15 20 25 30 35 40 D1 Forte Densité D2 D3 D4 Faible Densité HETRE*
SPADFigure 8 : Moyennes des masses surfaciques, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
Figure 9 : Moyennes des indices de teneur en chlorophylle, selon l’espèce, et en fonction du gradient de densité de tiges et du mélange d'espèces
11 moyennes sont peu différentes entre elles, à l’exception des arbres en monoculture et en forte densité (Fig.9).
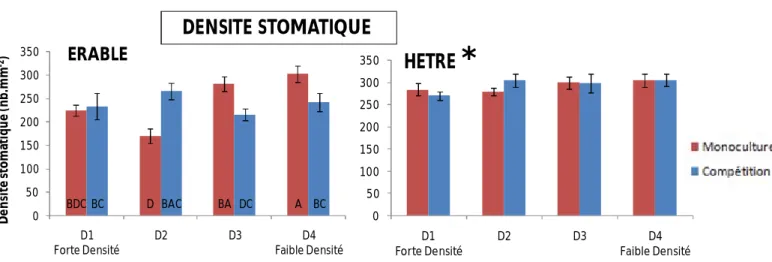
8) Densité stomatique
Les érables présentent des différences significatives de densité stomatique en fonction de la densité, mais en interactions avec le mélange (Tab.1) : en conditions de monoculture, le nombre de stomates a tendance à être supérieur dans les faibles densités, alors qu’en conditions de compétition interspécifique, les différences de densité stomatique entre les différentes classes de densité ne sont pas significatives (Fig.10). Les hêtres ne présentent pas de différences de densité stomatique quelque soit la densité de tiges ou les conditions de mélange (Tab.1).
IV.
DISCUSSION
Nous avons fait le choix d’analyser espèce par espèce les données obtenues, sans comparer directement le comportement des deux espèces étudiées.
1) Croissance et caractéristiques foliaires chez l’érable sycomore
Treize ans après la plantation, nous n’observons pas de très grandes différences de hauteur des érables entre les niveaux de mélange ou de densité de tiges (Fig.3), même si une légère tendance à de plus fortes hauteurs en compétition interspécifique et en faible densité a été obtenue. De plus, des plus faibles circonférences des arbres sont présentes dans les fortes densités de tiges (Fig.4). Cela semble indiquer, pour l’érable, une stratégie de faire des tiges plus élancées en conditions de forte densité, même si en fait cela ne se traduit pas par de plus fortes hauteurs pour ces arbres. L’absence de différences claires de hauteur entre les niveaux de densité pourrait
0 50 100 150 200 250 300 350 D1 Forte Densité D2 D3 D4 Faible Densité HETRE
*
BDCBC D BAC BA DC A BC 0 50 100 150 200 250 300 350 D1 Forte Densité D2 D3 D4 Faible Densité D e n si té s to m at iq u e (n b .m m -2) ERABLEDENSITE STOMATIQUE
12 s’expliquer par deux effets antagonistes de la compétition pour les ressources (en eau et éléments minéraux) et la lumière. En effet, dans les plus fortes densités de tiges, la quête de la lumière est plus importante, mais la faible disponibilité des ressources nutritives et lumineuse induit une croissance individuelle plus faible qu’on pourrait le supposer. La recherche de la lumière pour l’érable en forte densité semble être une stratégie privilégiée, mais la forte compétition entre les arbres pour les ressources (lumière, eau, éléments minéraux) ne leur permettraient finalement pas d’atteindre des hauteurs plus importantes qu’en faible densité.
Un résultat majeur de notre travail est que la compétition avec le hêtre engendre des différences de hauteur et de circonférence pour l’érable (Fig.3-4), même si, en fait, cela n’est réel que dans les faibles densités de tiges. Ces résultats montrent une évolution dans le comportement compétitif de l’érable. En effet, les conclusions de Vinkler et al. en 2007, ne montraient aucune influence de la compétition avec le hêtre sur la hauteur ou le diamètre de l’érable. Dans les fortes densités de tiges, la compétition pour les ressources est finalement aussi forte, que l’érable soit en compétition intra- ou interspécifique. Par contre, en faible densité, les ressources sont très disponibles, et il semblerait alors que la compétition pour ces ressources avec le hêtre stimule la croissance en hauteur et en diamètre des érables. Cet effet pourrait être lié aux différences d’architecture et de système racinaire entre les deux espèces, se traduisant par un effet réel dans les densités intermédiaires (c’est à dire D3). Néanmoins, cet effet est moins probable en très faibles densités (D4) car les houppiers ne sont pas en contact (donc pas d’effet de l’architecture), l’effet porterait alors éventuellement sur une compétition racinaire.
Les taux de croissance relatif sur la hauteur sur la période 2009-2011 ne diffèrent pas entre les différentes densités de tiges et l’effet compétition (Fig.5), alors qu’un effet de la densité sur le taux de croissance relatif de la surface terrière est apparu (Fig.6). En monoculture, c’est dans les faibles densités que ce taux est le plus fort, ce qui est cohérent avec l’effet densité observé sur la circonférence (Fig.4-6). Ces arbres, actuellement, semblent avoir une stratégie de renforcer leur circonférence (croissance secondaire), peut-être en lien avec l’effet du vent plus fort dans les faibles densités (Jaffe and Forbes, 1993). Le fait que ces arbres soient en compétition avec le hêtre ou d’autres érables ne semblent pas jouer.
Un résultat majeur de cette étude a été de mettre en évidence un effet significatif de la compétition entre espèces sur les caractéristiques foliaires de l’érable. Cet effet a été observé sur l’épaisseur des feuilles et la masse surfacique, alors qu’aucun effet n’a été observé pour l’indice de teneur en Chlorophylle (SPAD) et la densité des stomates (Tab.1). L’effet sur le LMA et l’épaisseur suggère un effet global de la compétition sur le fonctionnement des feuilles des arbres en compétition avec le hêtre, en particulier lié à la gestion du carbone foliaire. Nous avons observé
13 également un effet de la densité sur l’ensemble des caractéristiques foliaires mesurées (Tab.1), alors que nous n’avons échantillonné que des feuilles de lumière. Ce résultat souligne donc un effet direct de la compétition entre arbres sur les caractéristiques foliaires, qui d’ailleurs apparaît en interaction avec l’effet de la densité (Tab.1). Globalement, en fortes densités de tiges, les érables présentent des épaisseurs de feuille plus faibles quand ils sont en compétition avec le hêtre, alors qu’en faibles densités, il n’y a pas de différences significatives entre les niveaux de compétition (Fig.7). Le LMA diminue globalement dans les fortes densités, pour les deux niveaux de compétition, mais reste en général supérieur en conditions de mélange (Fig.8). Ces deux tendances sont cohérentes et suggèrent qu’en compétition avec les hêtres et en forte densité de tiges, les érables tendent à privilégier une stratégie de faire des feuilles plus étalées et fines, peut-être pour essayer de capter plus de lumière. Dans les faibles densités, l’érable ne semble pas subir cette limitation potentielle pour la lumière, quelque soit l’espèce avec laquelle il est en compétition, et nous n’observons pas de différences significatives d’épaisseur ou de LMA entre les différents niveaux de compétition.
L’effet de la densité et de la compétition sur la densité stomatique est complexe à analyser car l’interaction est significative (Tab.1). De plus, nous n’avons pas été en mesure d’estimer la taille des stomates. L’interprétation fonctionnelle des différences de densité stomatique, en termes de flux d’eau, entre les différentes situations de densité ou de compétition, est donc difficile, car des différences de densité stomatique pourraient être associées également à des différences de taille de stomates. De plus, nous pouvons souligner que l’estimation de la densité stomatique chez l’érable, au grossissement choisi, a été rendue difficile par la présence de nombreuses nervures tertiaires. Il nous a semblé que les valeurs obtenues pourraient ne pas être totalement représentatives des densités stomatiques réelles. Néanmoins, les différences de densité stomatique observées (Fig.10) confirment une influence forte de la densité de tiges et de la compétition entre espèces sur le fonctionnement foliaire de l’érable.
2) Croissance et caractéristiques foliaires chez le hêtre
En ce qui concerne les caractéristiques de croissance des hêtres, nous observons actuellement aucune influence de la densité de tiges ou du mélange d’espèces sur la hauteur des individus (Tab.1). Ce résultat est cohérent avec ceux obtenus en 2007 par I. Vinckler. Ce résultat peut s’expliquer en partie par le fait que le hêtre est une espèce relativement sciaphile dans les jeunes stades de croissance et avec un houppier très développé, présentant une architecture de type Troll (Oldemann, 1990). Dans les faibles densités, il profite ainsi largement de la lumière disponible pour sa croissance, alors que dans les fortes densités, la compétition avec les autres arbres, quelque soit l’espèce (hêtre ou érable), se traduit par une croissance finalement aussi forte qu’en faible densité, malgré une moindre disponibilité en lumière pour chaque individu.
14 Un résultat important de notre étude pour le hêtre, à la différence avec l’érable, est l’absence d’effet de la compétition interspécifique sur la hauteur des arbres après 11 ans (Tab.1 et Fig.3). De même, aucun effet significatif de la compétition pour le hêtre n’a été observé pour la circonférence ou les différents taux de croissance (Tab.1 et Fig.4-5-6). Le comportement du hêtre semble également avoir évolué, car d’après Vinckler et al., un effet du mélange apparaissait sur le diamètre des arbres en 2007. L’absence d’effet de la compétition interspécifique sur les caractéristiques de croissance du hêtre pourrait s’expliquer en partie par le fait que parmi les espèces feuillues tempérées d’Europe, le hêtre est une des espèces les plus compétitives, en tout cas en absence de stress hydrique (Backes & Leuschner 2000). Ainsi, dans la plantation étudiée, quelque soit l’espèce avec laquelle il est en compétition (hêtre ou érable), il semble maximiser sa croissance et aucun effet du mélange n’a été observé (Tab.1).
Un effet de la densité de tiges sur la circonférence des arbres a été observé (Tab.1), mais cet effet n’est visible en réalité qu’au niveau de la très faible densité des tiges (D4) (Fig.4). Comme pour l’érable, ce résultat peut être lié à l’effet du vent, plus fort dans les faibles densités.
Sur les quatre variables testées, seule l’épaisseur des feuilles a montré un effet significatif de la densité ou de la compétition (Tab.1). Ainsi, à l’exception de l’épaisseur des feuilles, les caractéristiques foliaires du hêtre, comme pour les caractéristiques de croissance, ne semblent pas être influencées par le mélange d’espèce, pour une densité de plantation donnée. Il est à noter néanmoins que l’absence observée d’effet de la compétition sur les caractéristiques fonctionnelles foliaires pourrait résulter de la conjugaison d’effets antagonistes sur les différents processus impliqués dans le fonctionnement foliaire, mais nous n’avons pas été en mesure d’approfondir ces effets. L’effet significatif de la compétition sur l’épaisseur n’apparaît par ailleurs que dans les faibles densités (D3 et D4) (Fig.7) : nous n’avons pas pu interpréter physiologiquement cette différence, étant donné que cet effet n’apparaît pas pour le LMA (Fig.8).
De la même manière que pour les érables, la taille des stomates n’a pas été mesurée. L’absence de différence de densité stomatique entre les niveaux de densité de tiges ou de compétition ne peut donc pas être directement interprétée d’un point de vue fonctionnel, associé à la transpiration foliaire. Néanmoins, l’absence de différences de caractéristiques foliaires confirme les observations sur la croissance du hêtre et le fait que les contraintes potentielles liées à la compétition pour les ressources entre des hêtres ou entre des hêtres et des érables n’influence pas son fonctionnement global.
15
V.
CONCLUSION
En conclusion, notre travail a montré que globalement, la compétition entre le hêtre et l’érable, pour une densité donnée, semble influencer le fonctionnement de l’érable, mais pas du hêtre. En conditions de compétition interspécifique, nous avons montré que la croissance de l’érable est globalement stimulée par le mélange d’espèces, alors que ce n’est pas le cas pour le hêtre. A contrario, pour le hêtre, l’effet de la compétition avec l’érable n’engendre pas d’effet négatif sur la croissance. Il apparait donc intéressant, pour le gestionnaire forestier, de mettre en plantation ces deux espèces en mélange.
Pour le hêtre, l’absence d’effet de la compétition interspécifique sur la croissance est cohérente avec l’absence d’effet sur les caractéristiques foliaires liées à l’acquisition des ressources (LMA, SPAD), même si l’épaisseur des feuilles semble être un peu supérieure en compétition, dans les faibles densités. Pour l’érable, l’effet de la compétition sur la croissance est associé à un effet sur les caractéristiques foliaires, même si cet effet apparaît principalement dans les fortes densités (à l’exception de la densité stomatique). Les différences observées sur la croissance des arbres semblent donc bien résulter de différences liées à l’acquisition des ressources (lumière, CO2,
éléments minéraux). Des mesures complémentaires de caractéristiques foliaires (teneur en N et C, composition isotopique du carbone et de l’oxygène) seront effectuées prochainement pour étudier plus précisément l’effet de la compétition entre espèces sur l’efficience d’utilisation de l’eau des arbres et la disponibilité en azote foliaire.
Nos données portant seulement sur des arbres qui sont installés dans la canopée, avec des feuilles de lumière, elles ne représentent pas l’influence globale de la densité ou de la compétition interspécifique sur le fonctionnement du peuplement. Il pourrait être ainsi intéressant de poursuivre ce travail en échantillonnant également des feuilles d’ombre de ces arbres.
BIBLIOGRAPHIE
Backes K., Leuschner C., 2000. Leaf water relations of competitive Fagus sylvatica and Quercus petraea
trees during 4 years differing in soil drought. Revue Canadienne de Recherche Forestière, Vol 30,
pp.335-346
Bock J., Richter C., Franc A., 2005. Panorama : Les forêts mélangées dans le nord de la France.
Rendez-vous Techniques, n°10 : pp. 36-42
Decocq G., Aubert M., Dupont F., Bardat J., Wattez-Franger A., Saguez R., De Foucault B., Alard D., Delelis-Dusillier A., 2005. Silviculture driven vegetation change in a European temperate deciduous
forest. Annals of Forest Science 62, pp. 313-323
Jaffe M-J., Forbes S.,1993. Thigmomorphogenesis : the effect of mechanical perturbation on plants. Plant
16
Landmann G., Landeau S., 2008. Les peuplements forestiers mélangés. Introduction aux ateliers
Recherche et Gestion Forestière (REGEFOR) 2007. IN Ateliers REGEFOR 2007 : Forêts mélangées :
quel scénario pour l’avenir ? Revue Forestière Française, LX2 : pp.99-105
Morneau F., Duprez C., Hervé J-C., 2008. Les forêts mélangées en France métropolitaine. Caractérisation
à partir des résultats de l’Inventaire Forestier National. IN Ateliers REGEFOR 2007 : Forêts
mélangées : quel scénario pour l’avenir ? Revue Forestière Française, LX2 : pp.107-120
Oldeman R.A.A., 1990. Forest : Elements of silvology. Berlin: Springer Verlag, p.624
Sardin T., Bock J., Becquey J., 2008. Les peuplements mélangés : enjeux et interrogations des
gestionnaires. IN Ateliers REGEFOR 2007 : Forêts mélangées : quel scénario pour l’avenir ? Revue Forestière Française, LX2 : pp.121-128
Vinkler I., Collet C. Ningre F., Faivre-Vuillin B., Bock J., 2007. Etude du mélange hêtre (Fagus
sylvatica)/ érable sycomore (Acer pseudoplatanus) : présentation d’un double clinal en forêt domanial de Haye (54). Présentation Atelier REGEFOR, 26-28 Juin 2007
17
ANNEXE 1 : ABREVIATIONS
- INRA : Institut National de Recherche Agronomique - LERFOB : Laboratoire d’Etudes des Ressources Forêt Bois - ONF : Office National des Forêts
- UMR : Unité Mixte de Recherche
- LMA : Leaf Mass Per Unit Area = Masse surfacique (g.m-2) - SPAD : Soil Plant Analysis Development
18
ANNEXE 2 :
· Comparaison de moyennes du LMA pour les érables, à l’aide de l’outil test sur les rangs multiples de Duncan sous SAS :
· Comparaison de moyennes du nombre de stomates pour les érables, à l’aide de l’outil test des rangs multiples de Duncan sous SAS :
19
ANNEXE 3 :
· Aperçu de la procédure GLM, pour la réalisation de l’analyse de variance multifactorielle, sous SAS :
20
RÉSUMÉ
De nombreux avantages potentiels sont conférés aux peuplements mélangés (Landmann et al. 2008). Cependant les mécanismes de fonctionnement de ces peuplements et les réponses des espèces lorsqu’elles sont en mélange sont encore mal connus. La meilleure conduite sylvicole à suivre dans ces peuplements n’est de ce fait toujours pas maîtrisée par les gestionnaires forestiers. L’objectif de cette étude est donc d’étudier l’influence de la densité de tiges ou du mélange d’espèces sur les caractéristiques de croissance et traits foliaires de deux espèces : le hêtre (Fagus sylvatica) et l’érable (Acer pseudoplatanus), dans le but d’améliorer ces connaissances. L’échantillonnage des arbres a été réalisé dans le dispositif double-clinal en forêt de Haye (54). Des mesures de hauteur, circonférence, épaisseur des feuilles, masse surfacique, indice de teneur en chlorophylle (SPAD) et de densité stomatique ont été effectuées sur 157 arbres. Les résultats obtenus montrent une influence du mélange sur l’érable. Ce dernier semble avoir sa croissance stimulée en condition de compétition interspécifique avec le hêtre. Concernant le hêtre aucun n’effet positif mais également négatif, de la compétition sur sa croissance, n’a été observé. A l’avenir, une étude complémentaire sera menée sur ce même dispositif afin d’étudier plus précisément l’effet de la compétition entres espèces sur l’efficience d’utilisation de l’eau des arbres et la disponibilité en azote foliaire.