Wide-range dispersal in juvenile Eagle Owls ( Bubo bubo ) across the European Alps calls for transnational conservation programmes
Texte intégral
Figure
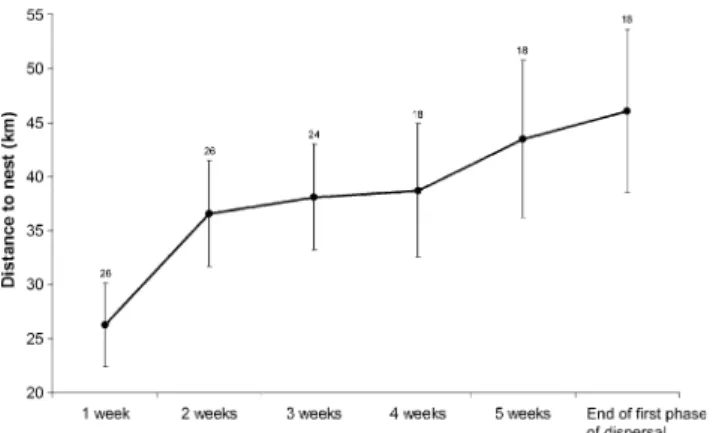
Documents relatifs
Il est vraisemblable que la date hâtive de ponte notée en 1988 ne cons- titue pas une exception (avec pour origine par exemple les conditions climatiques clémentes du début de
The results show that although mothers transfer their prey item to one of the offspring more rapidly than fathers once in their nestbox, nestlings begged more intensely in the
The analysis of a large and representative set of Malus x domestica genotypes indicated that apple germplasm diversity reflects its origination within three main geographic regions
Cela peut expli- quer que 85 % des bénéfi ciaires du CSP qui se sont inscrits à Pôle emploi en avril et mai 2014 restent inscrits sur les listes de demandeurs d’emploi du- rant
Error estimates for phaseless inverse scattering in the Born approximation at high energies.. Alexey Agaltsov,
Damage to Pigeon long bones in pellets of the Eagle Owl Bubo bubo and food remains of Peregrine Falcon Falco peregrinus: zooarchaeological implications. Stewart (Eds.), Proceedings
This swipe story supports an illustrated novel [2], and digital storytelling has been applied with the aim to bring the young audiences closer to the knowledge of the cultural
The best accuracy was obtained by using the VLAD approach that has been recently proposed for performing object recognition on a large scale1. Surprisingly, even the BoF approach
