Characterizing variation in mycorrhiza effect among diverse plant varieties
11
0
0
Texte intégral
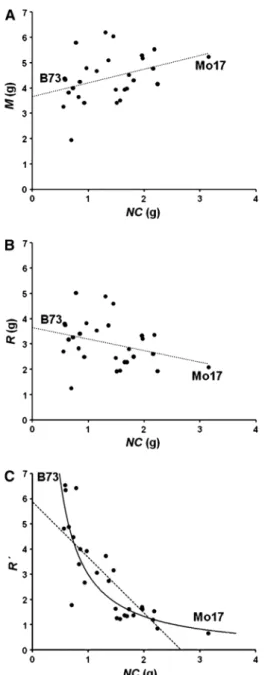
Figure
Documents relatifs