Artificial Gravity:
The Role of Visual Inputs in Adaptation to Short-Radius Centrifugation
by
Erika Lynn Brown
B.E. Biomedical Engineering
Vanderbilt University, 2000
SUBMITTED
IN PARTIAL
TO THE DEPARTMENT OF AERONAUTICS AND ASTRONAUTICS
FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE IN AERONAUTICS AND ASTRONAUTICS
AT THE
MASSACHUSETTS INSTITUTE OF TECHNOLOGY
JUNE 2002
0 2002 Massachusetts Institute of Technology
All rights reserved
AERO
LIARIS STTUTE OF TECHNOLOGYA
G 13 2002
LIBRARIES
Signature of Author:
Department of Aeronautics and Astronautics
May 22, 2002
Certified by:
Accepted by:
Laurence R. Young polo Program Professor of Astronautics Thesis Supervisor
Wallace E. Vander Velde Professor of Aeronautics and Astronautics Chair, Committee on Graduate Students
Artificial Gravity:
The Role of Visual Inputs in Adaptation to Short-Radius Centrifugation
By
Erika Lynn Brown
Submitted to the Department of Aeronautics and Astronautics
on May 23, 2002 in partial fulfillment of the
requirements for the Degree of
Master of Science in Aeronautics and Astronautics
ABSTRACT
Artificial gravity provides a critical enabling technology for long-duration human missions both within and beyond Low Earth Orbit by providing an integrated countermeasure to microgravity deconditioning. Unfortunately, the constraints of current vehicles necessitate the use of high-velocity short radius centrifuges and rule out large rotating spacecraft. Head turns made at these high rates of rotation introduce a host of problematic vestibular responses, including disorientating illusions of self-motion, improper reflexive eye movements, and motion sickness. Young, et al (2001) provided evidence that, unlike previously thought, the vestibular side effects of 23-rpm short radius centrifugation can be overcome with proper adaptation. The problem that remains is one of optimization.
We adapted 33 young adults to supine right-quadrant yaw head turns on-board a 2-m 23-rpm SRC. Data was collected both before and after a 10-minute adaptation period in one of three
visual conditions: DARK, STABLE with a centrifuge-fixed visual field, and EXTERNAL with
full view of the room. Four key results stand out:
1) Nearly all the parameters measuring subjective experience, including metrics of
motion sickness and illusory self-motion, decreased significantly across days and phases. The visual condition encountered during adaptation played no significant role
in this process.
2) There was a nearly significant effect of Day on the normalized slow phase eye velocity (NSPV) of (STABLE and EXTERNAL) subjects adapted with visual inputs. By contrast, no vestibulo-ocular reflex (VOR) adaptation was discernable for subjects adapted in the DARK. Visual inputs were required to adapt eye movements.
4) In subjects adapted with a view of the external world, the NSPV was significantly higher for turns to RED than to NU. This same trend was absent for groups adapted in the dark or with a stable visual field.
5) For all three experimental groups, head turns made to RED elicited less intense sensations of illusory motion than turns made to NU. This was in contrast to the differences described above for NSPV and points to dissociation between the adaptation of physiological parameters and subjective experience.
Thesis Supervisor: Dr. Laurence R. Young
Title: Apollo Program Professor of Astronautics Department of Aeronautics and Astronautics
This work was jointly supported by the National Space Biomedical Research Institute through a cooperative agreement with the National Aeronautics and Space Administration (NCC 9-58) and
June 2002
ACKNOWLEDGEMENTS
I would like acknowledge the Whitaker Foundation, National Space Biomedical Research
Institute, and National Space Society for providing the financial support for these dreams and believing in the value of education.
My sincere appreciation goes out to Dr. Laurence Young for his guidance and support on this
project and my other adventures here at MIT. Thank you for pointing me in the right direction and allowing me the freedom to dream larger.
Special thanks to Dr. Heiko Hecht for the constant input and intellectual debate that moved this research forward from day to day. Your unique perspective on the psychological aspects of the project and unfailing assistance with details were very helpful.
I would like to thank Nate Newby for all the long hours shared in classes, projects, and the lab.
Thank you for going the extra mile with all your hard work in programming, tweaking, and repairing that allowed the AG lab to flourish. I wish you, Nici, and Abigail all the best!
Many thanks to Dave Phillips for being for a fabulous officemate and friend. Your help in chugging through the slow days last spring was tremendous!
I owe much to Dr. Alan Natapoff for his innumerable nuggets of trivia, jewels of wisdom, and
insight into the enigmas of statistics. Alan, your patience and attention to detail are exceptional.
To Kathy Sienko, Lisette Lyne, Carol Cheung, David Balkwill, Peter Diamandis, and all the other pathfinders that came before me: thank you for blazing the trails for all of us to follow. This thesis owes much to your vision and persistence.
I am eternally indebted to the many UROPs who have offered their time, energy, exuberance, and
ideas to the project: Ethel Machi, Christopher Lee, Aaron Du, Dylan Chavez, Elliot Vasquez, Erin Alexander, Heather Knight, Alison Gallagher-Rogers, George Hanson, Lindsey Wolf, Jaclyn Cichon, Jason Herrera, Andrea Fanucci, and Rupal Parekh.
A big thank you to the rest of the MVL Staff and Crew for all the support you've provided
through nearly two years of research, quals, classes, and the wild world of MIT. The sunshine y'all have brought to the concrete castle has been irreplaceable.
My gratitude and love goes out to Greg, Lori, Paul, Carmen, Joaquin, and all the other friends that
have found time amidst our crazy schedules to share a bit of life here in Boston. Y'all have been a wonderful source of sanity!
Mom and Dad, thank you from the bottom of my heart for pointing towards the stars and encouraging me every step of the way. Your love and guidance has made all of this possible, and
I can't thank you enough.
Last but certainly not least, all my love and thanks to Andy, for being my rock and true companion throughout this whole process. Your unflagging support and encouragement have been more than any gal could ask for..
Erika L. Brown, S.M. Thesis 4
June 2002 TABLE OF CONTENTS 1. INTRODUCTION...9 1.1. Artificial Gravity ... 10 1.2. Adaptation ... 12 1.3. Sensory Conflict...13 1.4. Hypothesis ... 14 1.5. Outline...15 2. BACKGROUND... ... ... 17 2.1. Countermeasures...17 2.2. Vestibular Anatomy/Physiology... 20
2.3. Sensory Conflict in a Rotating Environment... 31
3. M ETHODS ... 35
3.1. Experimental Design... 35
3.2. Equipment...36
3.3. M etrics of Subjective Experience...38
3.4. Subjects...39
3.5. Protocol...40
3.6. Data Analy s ... 43
4. RESULTS ... 49
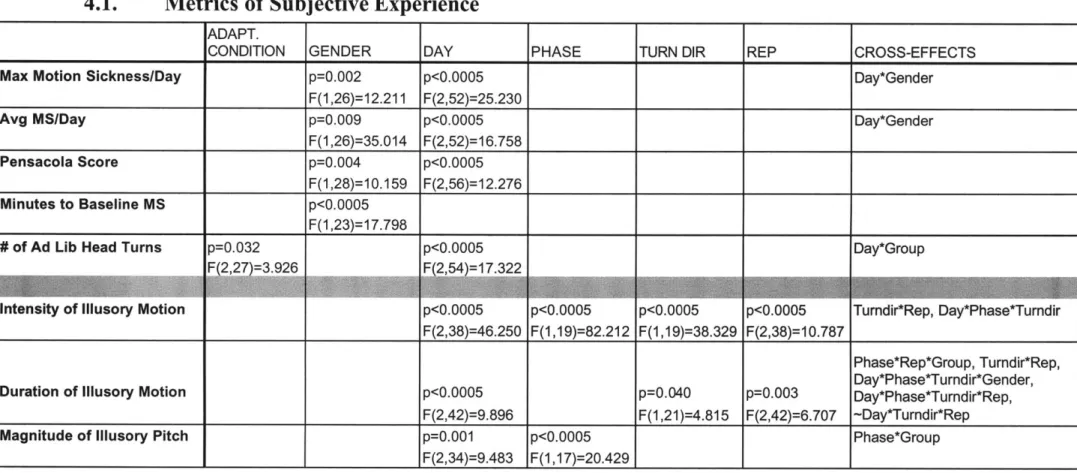
4.1. M etrics of Subjective Experience...50
4.2. Physiological M etrics... 58
5. DISCUSSION ... 65
5.1. Key Findings...66
5.2. Analysis...66
5.3. Operational Definitions of Adaptation... 72
5.4. Limitations & Recommendations for Future W ork... 75
6. CONCLUSIONS... o ... 79
REFERENCES ... ... ... ... .81
APPENDIX A - MODIFIED PENSACOLA MOTION SICKNESS RUBRIC (LYNE, 2000)...87
APPENDIX B - M EDICAL D ISQUALIFICATION LIST ... 91
APPENDIX C - INFORMED CONSENT FORM ... ... ... 92
APPENDIX D - PRE-TEST BRIEFING SCRIPT...94
APPENDIX E - EXPERIMENT PROTOCOL ... .. ... 96
APPENDIX F - POST-EXPERIMENT SURVEY... ... ... 101
APPENDIX G - INDIVIDUAL SUBJECT DATA...106
June 2002
LIST OF FIGURES
Figure 1: NASA 's first treadmill aboard Skylab ... 18
Figure 2: Anatomy of the vestibular system (Howard, 1986)...20
Figure 3: Detail of semicircular canal ampula (IMC, 2001)...21
Figure 4: Tip link transduction model of stereociliar gating (Pickles & Corey, 1992)...21
Figure 5: The fast and slow phases of a down-beating nystagmus ... 26
Figure 6: The MIT Man-Vehicle Lab 2-m short radius centrifuge with transparent canopy ... 36
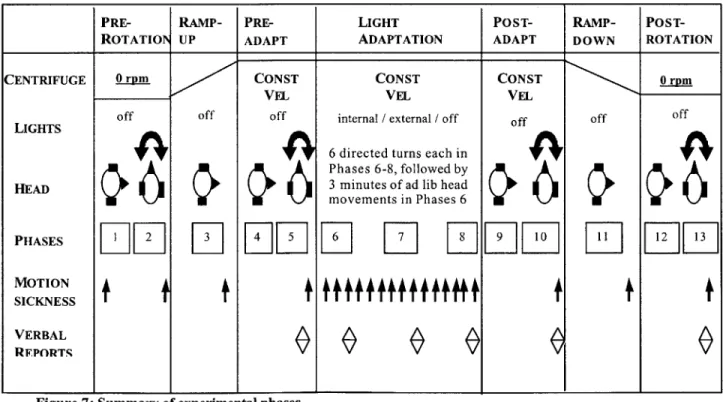
Figure 7: Summary of experimental phases... 41
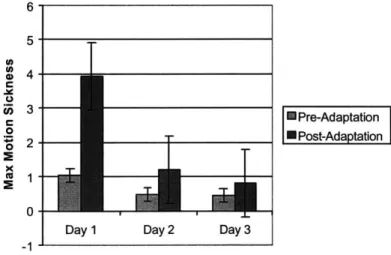
Figure 8: Maximum motion sickness scores by Phase and Day...51
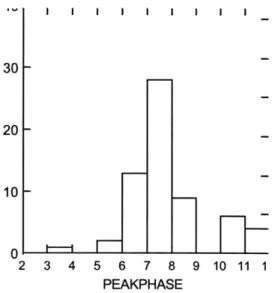
Figure 9: Histogram for Phase of peak motion sickness scores...52
Figure 10: Peak motion sickness scores by Gender and Day...52
Figure 11: Maximum Motion Sickness Scores per Day by Experimental Group. ... 53
Figure 12: Number of ad lib head turns made during adaptation periods (Phases 6 & 7) by Day and Experim ental Condition ... 54
Figure 13: Subjective Intensity Ratings of Illusory Self-Motion by Day, Phase, and Turn D irection ... 54
Figure 14: Pre- and Post-Adaptation Subjective Intensity by Day and Adaptive Visual Condition ... 55
Figure 15: Subjective Duration of Illusory Motion by Day, Phase, and Group...55
Figure 16: Magnitude of Illusory Pitch by Day, Phase, and Group...57
Figure 17: Magnitude of Illusory Pitch Sensations by Group and Phase...57
Figure 18: Histogram for Angle of Yaw Head Turns...59
Figure 19: Histogram for Time Constant, t, of Slow Phase Velocity Decay...60
Figure 20: Normalized Slow Phase Velocity measures across Day, Phase, and Adaptation Condition. Heavy lines indicate the trend from Day 1 pre-adaptation to Day 3 post-adap tation . ... 6 1 Figure 21: Student's two-sample t-test for head turns made to NU and RED by subjects adapted with a view of the external w orld. ... 62
Figure 22: Mean Friedman test rankings for NSPV by Adaptation Condition and Day...63
Figure 23: Standardized Normalized Slow Phase Velocity measures across Day, Phase, and A daptation C ondition...64
LIST OF TABLEs Table 1: Conflicting sensory modalities in each test group...35
Table 2: Average Subject Characteristics... 39
Table 3: Statistically significant (p<0.05) primary and cross effects on subjective metrics. ... 50
Table 4: Statistically significant (p<0.05) primary and cross effects on physiological metrics. ... 58
Table 5: Statistically significant (p<0.05) primary and cross effects on NSPV for each adaptation con dition . ... 62
Table 6: Two-sample t-test results for Normalized Slow Phase Velocity...63
Erika L. Brown, S.M. Thesis 6
June 2002
"YOU ARE NEVER GIVEN A WISH WITHOUT ALSO BEING GIVEN THE POWER TO MAKE IT COME TRUE. YOU MAY HAVE TO WORK FOR IT, HOWEVER."
RICHARD BACH
"ANCORA IMPARO"
"I AM STILL LEARNING"
MICHELANGELO BUONARROTI
"THERE WAS A STAR DANCED, AND UNDER THAT WAS I BORN." WILLIAM SHAKESPEARE, TWELFTH NIGHT
"AND THOUGH I OFT HAVE PASSED THEM BY,
A
DAY WILL COME AT LAST WHEN I SHALL TAKE THE HIDDEN PATHS THAT RUN WEST OF THE MOON, EAST OF THE SUN."J.R.R.
TOLKEIN"THE URGE TO EXPLORE IS A FUNDAMENTAL HUMAN IMPULSF, WHICH NEEDS NO FURTHER JUSTIFICATION... AND WE WANT TO PLAY WITH SPACESHIPS." ARTHUR C. CLARKE
Erika L. Brown, S.M. Thesis 7
June 2002
Erika L. Brown, S.M. Thesis 8
June 2002
1.
INTRODUCTION
The entire process of biological evolution has taken place in the constant presence of Earth's gravitational force. However, in the last forty years, the age of space travel has placed unique demands on the body, as every physiological system has been asked to deal with the novel weightless environment of orbit. When these challenged systems are compared with their ground-based steady-state, various deficiencies are found.
In the absence of the steady influence of gravity, fluids shift from the lower body up towards the head and torso, causing facial puffiness and an increased load on the cardiovascular system. To return the body to homeostasis, the renal system begins a process of diuresis and fluid shedding. This in turn causes the hematocrit of the blood to rise, which leads to a reduction in erythropoesis, and to lysis of existing red blood cells.
In response to decreased loads, the osteoblasts of the skeletal system reduce production of new bone, leading to an imbalance with osteoclastic activity and an overall reduction in bone mineral density. Muscles, particularly the extensors of the lower body, atrophy from disuse, and show a preferential shift from oxidative slow twitch fibers, which provide endurance and long-duration posture control, to glycolytic fast twitch fibers, which provide precision control and short bursts of strength. The cardiovascular system no longer has to respond to the hydrostatic gradients produced by changes in body orientation on Earth, so there is a down-regulation of the baroreceptors and an atrophy of the cardiac muscles (Diamandis; NRC, 1998, 2000).
In addition, the neurovestibular mechanisms that regulate balance and motor control are faced with a novel environment, devoid of many of the usual cues used for coordination. Thus, the brain learns to rely preferentially on vision for orientation, rather than on signals from the visual and somatosensory systems. Otolith signals of linear acceleration are no longer interpreted as tilt of the head relative to gravity, but instead as pure linear translations. Strategies of motor control must also be adjusted to respond to the lack of normal gravity (Lathan & Clement, 1997; Young
& Sinha, 1998).
While the changes mentioned above are completely appropriate to the novel environment of space, current astronauts and cosmonauts are in that environment only as visitors, and not as residents. The greatest problems arise when they must return to a gravitational environment, be it lunar, martian, terrestrial, or otherwise.
June 2002
Although the current era of low-earth-orbit flights places spacefarers within close reach of the medical resources of Earth, future flights will take us beyond the horizon of immediate help. And as mission durations stretch beyond "months" into the domain of "years," the issues of readaptation will become even more salient.
In particular, as human spaceflight moves farther afield, the need for a comprehensive suite of countermeasures becomes more and more critical. The current cadre of options includes exercise, lower-body negative pressure (LBNP), fluid loading, and pharmacologic measures. However, while each of these addresses the symptoms of adaptive decline, none of them address the root of the problem itself, and no combination has proven adequate for long-duration spaceflight
(Cavanagh, Davis, & Miller, 1992; EDOMP, 1999; Goodship, et al, 1998; Nicogossian, Rummel,
Leveton, & Teeter, 1992). Artificial gravity, on the other hand, uses the centripetal accelerations of a rotating environment to directly counter the problems of microgravity. Rather than targeting one physiological system or symptom at a time, by synthesizing gravity iself, AG offers an integrated countermeasure for combating skeletal losses, muscular atrophy, cardiovascular deconditioning, and neurovestibular adaptation.
1.1.
Artificial Gravity
The traditional AG paradigm of a large-radius rotating spacecraft offers an ideal solution to microgravity deconditioning. As put forth by Verne, Tsiolkovsky, Von Braun and others, a single torus or tethered vehicle could be set in motion at a very low angular velocity to produce the desired gravitational forces in living and working quarters (Young, 1999). Stone's review (1970) of the human performance issues surrounding artificial gravity pointed to an ideal radius between
15.2 and 16.8 meters. Unfortunately, the construction and operation of such a vehicle raises
substantial political and engineering barriers. Although these large-radius gravitational systems are not practical in the near future, onboard short-radius centrifuges (SRCs) may offer a reasonable low-cost alternative.
By providing periodic "doses of gravity" to crewmembers aboard a standard spacecraft, SRCs
would offer an intermittent countermeasure to microgravity deconditioning. A number of ground studies have provided evidence that short-radius centrifugation does indeed stimulate the baroreceptors, and may thereby provide in-flight "training" for maintenance of a proper orthostatic response upon return to a gravitational environment (Iwasaki, 2001; Burton & Meeker,
June 2002
1992). Musculoskeletal atrophy may also be mediated by artificial gravity (D'Aunno, Thomason, & Booth, 1990) particularly if the static rotational stimulus is matched with the dynamic loads of
exercise (Gurovsky, et al, 1980).
Unfortunately, these SRCs are not without their own problems. As given by the basic physics equation, ac=or, when the radius (r) of a centrifuge is decreased, the rate of rotation (W) must be increased to maintain a given level of centripetal acceleration (ac). Therefore, while a 45-m radius rotating spacecraft would only need an angular velocity of 4.5-rpm to produce 1-g (9.8-m/s2) at its rim, a 1.7-m radius SRC would have to rotate at 23-rpm to produce the same "gravitational" force.
By the same token, for a given rate of rotation, the acceleration level will vary radially from hub
to rim, producing a gravity gradient along the radius of the spacecraft. For a 1.7-m (h) tall human standing inside the outer rim of a 45-m radius (r) spacecraft, rotating so as to produce l-g (ac) at the feet, a gravitational level of 0.96-g (ac(r-h) / r) would be felt at the top of the head, producing a 3.7% gravitational gradient down the astronaut's body length. However, on a 2-m radius SRC, this same individual would experience a 100% gravity gradient from head to toe. While studies have shown tolerance of Air Force pilots to high-G level 100% gradients (Piemme, Hyde, McCally, & Potor, 1986), no studies have been undertaken to examine the long-term effects of intermittent gradient exposures.
Perhaps most notably, however, head tums made at the high rotational velocities required for short-radius centrifugation introduce a host of problematic vestibular responses. When an astronaut or subject in a rotating environment makes head turns relative to the plane of rotation, the endolymph of the semicircular canals undergoes a cross-coupled acceleration equal to
-((Oentrifuge x ('lead), and experiences a torque, therefore, that is perpendicular to the motion both of
the head and of the centrifuge itself. While this cross-coupling makes a difference during the course of a head turn, its effect dies out very quickly after the turn is completed.
The changes in angular velocity caused by bringing the semicircular canals into or out of the plane of rotation, however, are much longer lasting, with a dominant time constant on the order of
6 seconds (Goldberg & Fernandez, 1971). These unusual changes in rotational equilibrium can
cause disorientating sensations of illusory self-motion, improper reflexive eye movements, and motion sickness.
June 2002
Recent work in the MIT Man-Vehicle Lab has shown promise that these effects, while disorienting and often nauseating, may be transient if a proper adaptation schedule is followed on board the SRC. Work by Young, Hecht, Lyne, Sienko, Cheung, and Kavelaars (2001) demonstrated an attenuation of illusory tilt, inappropriate vertical nystagmus, and motion sickness over the course of a one-hour 23-rpm centrifugation, with further adaptation over multiple days.
1.2.
Adaptation
The term "adaptation" has been used in a number of different connotations to describe changes to sensory afferents or their resulting responses during repeated stimulation. However, in many ways, this process is best described as two separate levels of change. First, repeated or prolonged application of a given stimulus may induce a simple decrease in response, known as "habituation." As exemplified by an individual who becomes accustomed to a persistent background noise or scent, after repeated signals, the body habituates and ceases to respond at the same level to the "old" stimulus as it would to a "new" one.
However, while habituation is quite common, it serves only to make the system less responsive to stimuli that are presented repeatedly. True adaptation, on the other hand, refers to changes that serve a more functional purpose. Perhaps one of the most extreme examples of adaptation is seen in the flounder, or flatfish. As it matures, the flounder changes from a vertical free-swimmer with one eye on each side of its head, to a bottom-dweller with both eyes on its dorsal side. This migration of one eye across the head to accommodate the new configuration necessitates significant changes in the neural interpretation of images and orientation. In order to provide accurate vision and coordination, the fish must adapt (Graf & Baker, 1985).
In the case of centrifugation, the improper reflexive eye movements that occur in response to head turns cause images to shift or "slip" across the retina. The accompanying sensations of illusory self-motion are detrimental to balance and orientation, and concomitant motion sickness provides a barrier to normal daily activity. Habituation causes a slight reduction in the magnitude of these responses across the period of the protocol on one day, but adaptation is needed to enable persistent reinterpretation of the sensory signals in such a way as to provide a useful input to the maintenance of visual stability and orientation, without causing disruptive nausea.
Erika L. Brown, SM. Thesis
12
June 2002
This adaptation also possesses the additional property of being extremely context-specific. While a subject is adapting on a rotating platform, nystagmus and illusory self-motion are appropriately reduced. Once rotation ceases, however, there is no carryover of inappropriate eye movements into the stable environment. Young, et al (2001) showed that, in fact, there were virtually no after-effects following 20-30 minute centrifugation sessions on three consecutive days. Some feedback or feed-forward mechanism must exist to provide a "trigger" to the system, indicating which paradigm is most appropriate for each context it is presented.
1.3.
Sensory Conflict
The Sensory-Conflict Model described by Reason (1978), Oman (1996), and others states that motion sickness onset may be initiated by a conflict between sensory input and the expected
sensations as determined by prior experience. Persistent conflict leads to an updated model and adaptation to the stimulus. More generally, Dichgans and Brandt (1978), and Bles (1981) applied the idea of a conflict model to inter-sensory disagreements between multiple end organ or
cognitive inputs. They showed that stronger inter-sensory conflicts drive stronger illusory sensations of self-motion.
In the case of a rotating centrifuge, there are a number of potentially conflicting stimuli,
including:
* Vestibular (Otolith and Semicircular Canals) * Visual * Tactile * Kinesthetic * Proprioceptive * Visceral * Auditory * Cognitive
* Motor Efferent Copy
Erika L. Brown, S.M. Thesis
13
June 2002
In keeping with the Sensory-Conflict Model and the theories of inter-sensory interaction above, it would seem that one could speed adaptation to a stimulus by maximizing the conflict between opposing sensory inputs. We term this the Conflict Theory of Adaptation.
For a blindfolded subject lying supinely on a centrifuge, rotating at constant velocity with her head on axis, there are very few sensory inputs that signal rotation. In fact, as long as her head is kept stationary to maintain vestibular equilibrium, the only rotational cues are provided by ambient sounds of the centrifuge, tactile wind cues, cognitive knowledge, and visual inputs. While each of the pieces of information plays a role in determining the total level of conflict, the visual-vestibular conflict is the most salient. Thus, this thesis examines the Conflict Theory of Adaptation for varying levels of visual-vestibular discrepancy.
1.4.
Hypothesis
Specifically, we hypothesize that, utilizing a stimulus of repeated yaw head turns in a supine rotating environment:
Maximizing the conflict between visual and vestibular inputs will result in more
intense subjective reactions and better adaptation to 23-rpm short-radius
centrifugation, as measured by reductions in motion sickness scores, inappropriate
vertical nystagmus, and magnitude and duration of illusory tilt.
Among subjects presented with a stable visual world (STABLE), a rotating view of the external visual world (EXTERNAL), or a darkened visual field (DARK), we expect that the STABLE condition, with rotating vestibular inputs and stable visual inputs, will provide maximal conflict, and thereby will produce both the largest initial reactions to the stimulus and maximal adaptation over the course of the experiment. We recognize that this runs counter to the concept of incremental adaptation (Graybiel, Deane, & Colehour, 1969; Lackner & Lobovits, 1978), which aims to speed adaptation by gently increasing the rate of rotation at a level below the threshold for motion sickness. However, if our higher initial level of conflict is tolerable, then a more rapid adaptation is certainly advantageous.
Erik L. rown S.M Theis 1
June 2002
1.5.
Outline
Chapter 2 provides the background motivation for this work, along with a detailed look at the
physiology of the vestibular system and its interactions with the visual pathways. Various models of sensory conflict are presented and a discussion of the term "adaptation," as used in this
context, follows.
Chapter 3 describes the experimental design used in this study, including subject selection,
hardware, data collection, and protocol. The methods used for data analysis are explored and some limitations are considered.
Chapter 4 presents the results of the study. The outcomes of several repeated measures analyses
of variance (ANOVAs) are presented, along with other statistical and numerical outcomes.
Chapter 5 discusses the significance of these results and their place in the broader body of
adaptation literature. The nature of differential adaptation between physiological parameters and subjective experience is considered, and the operational definition of "full adaptation" is explored. Limitations of the research are discussed and recommendations are presented for future explorations in the domain of short-radius centrifugation.
Chapter 6 summarizes the most critical results of this experiment and offers some conclusions to
the research.
Erika L. Brown, S.M. Thesis
15 15 Erika L. Brown, S.M. Thesis
June 2002
Erika L. Brown, S.M. Thesis 16
June 2002
2.
BACKGROUND
2.1.
Countermeasures
Over the 40-year history of human spaceflight, the suite of countermeasures to prevent microgravity deconditioning has evolved and grown on the basis of both research and preference. The earliest of flights of both the Russian and American space programs relied almost entirely on selection of healthy crew members and extensive training for high-g, high-stress environments. Over time, however, the missions changed from short duration exhibitions of capability to longer stays in microgravity geared more towards scientific experimentation, repair and construction of structures in orbit, and demonstrations of technologies for future missions. This paradigm shift has necessitated a change in the way countermeasures are viewed, and current efforts may be classified under the headings of Selection, Training, Lifestyle, Exercise, Fluid Redistribution, and Pharmacology.
The first step toward preventing extreme deconditioning in microgravity is the judicious selection of astronaut candidates with a low pre-existing susceptibility to such problems. By selecting healthy crew members with no serious history of medical or psychological difficulties, NASA and the Russian Space Agency head off many potential issues at the pass. Unfortunately, in the case of space motion sickness, no adequate measure has been found to predict susceptibility pre-flight.
Training provides another proactive countermeasure by offering astronauts and cosmonauts strategies for coping with the symptoms of deconditioning. The Russian Institute for Biomedical Problems utilizes extreme centrifugation training to try to accustom cosmonauts to the severe sensory conflict they may experience on orbit. Pre-flight adaptation trainers (PATs) have also been proposed to familiarize astronauts with the sensory rearrangements they may experience in-flight in order to mitigate space motion sickness (Parker, et al, 1985; Reschke, et al, 1988).
Countermeasures that fall under the heading of Lifestyle are aimed at maintaining a level of physical and emotional health. These include regular communication with family and friends on the ground, sufficient personal time, dedicated sleeping time, appropriate hygiene, and a healthy diet. Dietary deficiencies, in particular, have long been known to cause medical and psychological problems. For centuries, sailors on long sea voyages without fresh fruits and
June 2002
vegetables often suffered from tender gums, reopening of wounds, and the general deterioration
of scurvy. While citrus fruits were known to be a useful remedy, it was not until the
20' century
that a lack of vitamin C was incriminated as the cause of the disease. Insufficient calcium intake
has been implicated as a prime player in development of osteoporosis, and Caillot-Augusseau, et
al (2000) recently discovered that vitamin K supplements may help prevent bone loss during long
space flights.
Although diet alone cannot prevent deconditioning, careful regulation before,
during, and after flight can at least help to mitigate the inherent risk.
However, while careful Seection, Training, and Lifestyle decisions are helpful, they are not
enough to completely resolve the questions of deconditioning. To date, exercise has offered the
most obvious countermeasure to the remaining effects of spaceflight. In fact, current astronaut
crews on board the International Space Station are scheduled for a minimum of two hours of
exercise a day throughout the length of their stay. The Russian Space Agency focuses on an
intense exercise regimen near the end of the mission in an attempt to prepare the body for its
return to Earth.
Treadmills, which started on Skylab 4 as a Teflon-coated titanium plate and a belt with bungees
tied into the floor (Figure 1), have now taken on the elaborately damped form of the Treadmill
Vibration Isolation System, TVIS. Cycle and rowing ergometers have scaled down from their
early counterparts to become smaller, lighter, and more comfortable. Resistive exercises, such as
physical therapy bands, "Penguin Suits," and the spring-loaded Mini-Gym, have added variety to
the strength-building regimens (de Hon, 1997). However, while all of these systems provide
muscular loading and the first three provide the added bonus of aerobic cardiovascular exercise,
none provides the impact loading needed for high
levels of osteogenesis, none provide significant
vestibular stimulation, and none directly address
the extensive fluid shift seen in microgravity.
Three techniques that do address the issues of fluid
redistribution and volumetric loss are fluid loading
prior to reentry, anti-g suits, and Lower Body
Negative Pressure (LBNP). In the first two cases,
hydrostatic support is given to the body to help
Figure 1: NASA's first treadmill aboard Skylab prevent syncope upon return to gravity. LBNP,
June 2002
however, is unique among these options in that it can be used effectively during flight to provide both transient fluid redistribution and baroreceptor conditioning. Unfortunately, the loads placed
by LBNP on an already-weakened cardiovascular system may increase the risk of syncope.
Combining LBNP with dynamic leg exercise can help to mitigate this risk (Watenpaugh, et al,
1991).
Pharmacologic interventions offer a final barrier against deconditioning by acting either directly at the root of the problem, or at very least, ameliorating the symptoms. Promethazine has been used for years to minimize the problems of space motion sickness. The Shuttle Orbiter Medical System (SOMS) stocks numerous medications for symptoms ranging from congestion and allergies to sleeplessness and pain. The oral contraceptives taken by many female astronauts may offer some protection against bone demineralization (Harm, et al, 2001), and the Russian program has used medications, inclhding phenoxybenzamine and papaverine, to improve regulation of blood circulation (Martin, 1999). Many other treatments, particularly for prevention of musculoskeletal atrophy, are under investigation in human and animal models. Unfortunately, pharmacokinetics and pharmacodynamics, the distribution and utilization of drugs by the body, may be significantly altered by microgravity, which makes validation of these countermeasures quite difficult (Putcha & Cintron, 1991).
Unlike most of the strategies discussed above, intermittent artificial gravity offers an integrated approach to preventing deconditioning throughout the body. The advantages of SRCs alone are purely static. They offer little more maintenance value than standing upright against a wall on Earth. However, the advantages of even this minimal loading are great. Using a 6" head-down bed rest study to mimic the effects of microgravity exposure, Vernikos and her team (1996) showed that standing for 4 hours a day completely prevented losses in orthostatic tolerance and plasma volume reduction. The 18.5-day flight of Cosmos-936 rotated 10 Wistar rats at 53.5-rpm to produce 1-g of centripetal acceleration. Post-flight analysis of these animals and their microgravity control counterparts showed that AG provided a level of protection against otolith deconditioning, decreased stress responses, fewer metabolic disorders, improved immune characteristics, improved physical endurance, and decreased skeletal calcium loss. Problems arose, as described earlier, with unusual stimulation of the semicircular canals, as demonstrated
by reduced sensitivity to angular accelerations (Gurovsky, et al, 1980).
Erika L. Brown, S.M. Thesis
19
June 2002
By coupling artificial gravity with exercise systems, the advantages of dynamic loading in an
accelerated environment can be added to the static effects described above. Kreitenberg's SpaceCycleTM, developed by Cook Brothers Cycling, and the Ames Research Center's Human Powered Centrifuge (HPC) both offer the exercise advantages of a cycle ergometer coupled with centrifugal physiological benefits. Kreitenberg's 1995 SpaceCycleTM patent even offers a spring-loaded cam and roller device integrated into the pedals to provide "impact loading of normal earth walking." Preliminary evidence with the HPC suggests that exercise with +Gz acceleration (along the long axis of the body) does indeed add significant physiological stress as compared with passive rotation. At accelerations ranging from 1-g to 4-g, mean heart rate in exercising subjects was elevated 43-50 beats per minute above that of passively rotating subjects (Greenleaf,
et al, 1996).
Before we can even consider such a comprehensive countermeasure for spaceflight, however, a number of issues must be resolved, particularly those relating to the physiology of the neurovestibular system.
2.2.
Vestibular Anatomy/Physiology
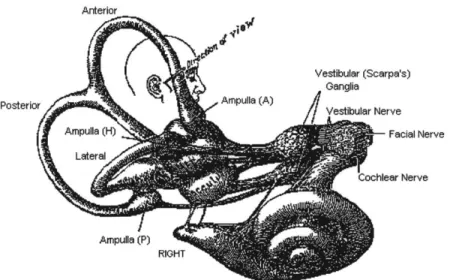
The human vestibular system provides data on linear and rotational accelerations as an input to the body's orientation, perception, and postural control systems. Positioned within the inner ear on each side of the head, the semicircular canals and otolith organs serve as physiological transducers, translating physical motion into the chemo-electric language of neural impulses
(Sherwood, 1997)(Figure 2).
Anterior
Vestibular (Scarpa's) Ganglia
Posterior Ampulla (A) Vestibular Nerve
Ampulla (H) Facial Nerve
Lateral
Cochlear Nerve
Figure 2: Anatomy of the vestibular system (Howard, 1986).
Erika L. Brown, S.M. Thesis 20
June 2002
Semicircular Canals
Rotations are sensed by a set of three orthogonal semicircular
canals in the inner ear. One set of these lateral, anterior, and
posterior canals is labeled in Figure 2. While the canals are
turned outwards approximately
35*-65*
from the medio-lateral
plane, and tilted upward 15*-30* from Earth-horizontal, theeffects may be described in terms of their components in the
body's roll, pitch, and yaw planes. It is worth noting, however,
s
of
hair cellethat the anatomical variability between subjects does introduce
some discrepancy in the actual components contributed by each
canal and could help to explain some differences in individual
Ampu"aCrftaresponses to rotational stimuli (Stone & Letko, 1965).
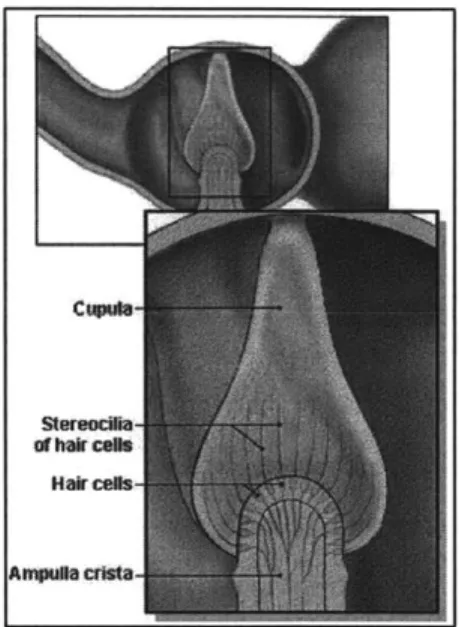
Figure 3: Detail of semicircular canal ampula (IMC, 2001).
_ _ Each of the semicircular canals consists of a membranous tube,
cushioned within the temporal bone by a viscous perilymph fluid,
and filled with another fluid known as endolymph. As seen in
)
I
Figure 3,
the base of each semicircular canal widens out into a
bulbous ampulla. Forming a seal across this ampulla is a gelatinous
structure known as the cupula, which in turn supports a series of
J
innervated hair cells. Each hair cell includes a number of microvilli
projections, organized with one larger kinocilium situated next to
20 to
50
shorter stereocilia. Rotations of the head in the pitch, roll,
Figure 4: Tip linkand yaw planes cause the endolymph in the
plane of rotation to transduction model of move with respect to the walls of the canals. The inertial lag of this stereociliar gating(Pickles & Corey, 1992) fluid causes the cupula to deform transiently in the direction opposite the head motion, thus bending the microvilli projections inside. As demonstrated in Figure 4, when the stereocilia are bent towards the kinocilium of a hair cell, thin filamentous tip links connecting the smaller hairs are stretched, opening an ion channel. A rapid influx of alkali cations, most likely K and Ca**, depolarizes the cell. In the opposite case, as stereocilia are bent away from their respective kinocilium, the gates to the ion channels are closed, and the hair cell becomes hyperpolarized (Pickles & Corey, 1992). These
chemoelectrical signals are consolidated by the vestibular nerve causing an analogous variation in
Erika L. Brown, S.M. Thesis 21
June 2002
nerve firing rates such that depolarization increases the rate of afferent signaling from the resting discharge and hyperpolarization decreases the firing rate (Sherwood, 1997).
In 1931, Steinhausen first described the fluid dynamics of the semicircular canals as analogous to an overdamped torsion-pendulum with an elastic restoring force (Equation 1):
Ea(t)
=
OE
(t) + fl (t) + A (t)
(1)
The torsional input of an angular acceleration of the head, x(t), is balanced by the angular acceleration of the cupula,
E
d2 (t)/dt2; the viscous drag of the endolymph, H d (t)/dt; and the cupular elastic restoring force, A (t); where H represents the damping frictional drag of the endolymph fluid, A is the spring constant of the cupular restoring force,E
is the moment of inertia of the toroidal fluid ring, and (t) is the angular displacement of the cupula with respect to time.The system is assumed to be second-order overdamped with two distinct real roots. Thus, by setting initial conditions to zero, taking the transformation into the Laplace domain, and collecting like terms, the model takes on a classic transfer function form (Equation 2), where s is the Laplace variable. The dynamics of this system are governed by both a long time constant,
,r= H/A, and a shorter time constant, T2 =EIH:
=/A
(s+A/H
)(s+H/O)
Van Egmond, Groen, and Jongkees (1949) provided the first theoretical and experimental estimates for the values of these time constants in humans, with a long time constant, 1 , around
10 seconds, and a shorter time constant, T2, of approximately 1 second. These numbers, however,
relied on inaccurate estimations of the radius of the semicircular canals, as well as measurements taken at the level of nystagmus and subjective sensations. By utilizing these downstream
measures, a large component of central processing was also included in the estimations and true estimations of the end organ time constants became impossible.
Erika L. Brown, S.M. Thesis
22
June 2002
Later work by Goldberg and Fernandez (1971) used direct neural monitoring in squirrel monkeys to circumvent these confounding effects and found time constants of 5.7 and 0.003 seconds, respectively. Asymmetries were seen between excitatory and inhibitory responses, with excitation causing larger deviations from the resting discharge. Goldberg and Fernandez posited that this asymmetry helps to prevent the nonlinearity of neurons being silenced, or driven down to a discharge rate of zero, in response to large inhibitory stimuli. The work also identified a high frequency lead component, hypothesized to provide compensation for the delays and phase lags introduced by the vestibular reflex pathways. And perhaps most notably, Goldberg and Fernandez confirmed an "adaptation" response modeled earlier by Young and Oman (1969). This adaptation was evidenced in some units by a decreased gain in response to prolonged stimuli, and a brief period of secondary response reversal following the end of acceleration. Addition of both lead and adaptation terms updated Steinhausen's original model to a more accurate form:
TAS (+TLS)
(1
+TAS)(l
TIS)(1
+t
2S)The frequency response curve for the model detailed above indicates different response patterns at different stimulus frequencies. For stimuli between about 0.1 and 10.0-Hz, initial cupular displacement, and thus canal output, provides accurate information about head angular acceleration. However, at frequencies of less than 0.1-Hz, the sensitivity of the canal response declines and a phase lead component becomes evident. At these low speeds, accurate information about angular motion is provided predominantly by visual inputs (Waespe & Henn, 1977; Paige, 1983).
Fernandez and Goldberg (1971) estimated the value of t1 based on its contributions to low
frequency phase lag in neuronal units with little evidence of adaptation response:
T1= (1/27rf) tan(4)
(4)The "convenient" low frequency, f, was chosen at 0.025-Hz, and an average phase lag, $, was obtained by direct measurements from 12 Low Adaptation (LA) units.
Erika L. Brown, S.M. Thesis 23
June 2002
12 was estimated by hydrodynamic calculations, utilizing values from Igarashi (1967) for the
internal radius of the squirrel monkey semicircular canal.
The high frequency lead and adaptation time constants, TL and TA, were estimated from
discrepancies, A$, between Steinhausen's original model and measured responses:
TL=
(1/2rf) tan(A$)
(5)
TA=
(1/27tf) tan(n/2
-
AO)
(6)
High frequency discrepancy for TL was calculated as an average of all the direct measurements available for sinusoidal stimulations between 1.0 and 8.0-Hz. For calculating TA, another "convenient" low frequency, f, was chosen - this time at 0.0125-Hz - and discrepancy was obtained by direct measurement. Average values in the squirrel monkey were found to be TL = 0.049 sec, and TA = 80 sec.
While direct measurement of these values in humans is impractical, modeling efforts have found values for the longer time constants, T, and T A, to be comparable with those found in monkeys.
While differences in physical dimensions of the semicircular canals would be expected to drive changes in t2, the value for this short time constant is so small as to be virtually irrelevant for the system in question.
Otolith Organs
Linear accelerations, including those of a gravitational field, are transduced by the saclike otolith organs. Also nestled within a cavity in the temporal bone, between the semicircular canals and the cochlea, the utricle and saccule are likewise filled with endolymph and cushioned by he viscous perilymph. Each of the organs consists of a curved macular plate composed of a number of hair cells (very similar to those of the semicircular canals) projecting into a gelatinous sheet. Embedded in the top layer of this matrix are small calcium carbonate "stones" which serve to raise the specific gravity of the membrane with respect to the surrounding endolymph (Graybiel,
1973). In a physiological response very similar to those described above, linear accelerations
cause sagging of the gelatinous sheet in the respective otolith organ. This sag in turn displaces
June 2002
hair cells, causing membrane depolarization or hyperpolarization, which triggers a change in the firing rate of afferent nerve impulses.
By treating the embedded calcium carbonate otoconia as a seismic mass with linear spring
restoring forces provided by the support structures of the macula, and inertial drag due to the viscous endolymph that fills the saccules and utricles, the otolith organs may be modeled analogously to the semicircular canal paradigms described above. The "effective mass" of this system, m, includes the endolymph that is carried along by the movement of the macula, such that:
ma(t)=
(Pe /
Pm)mX(t)
+
IIx(t)
+
kx
(t)
(7)The input of a linear acceleration of the head relative to inertial space, ma(t), is balanced by the linear acceleration of the buoyant otoconial mass with density, p, in endolymph of density, pe,
(Pe /p)m d2x(t)/dt2; the viscous drag of the endolymph, II dx(t)/dt ; and the macular elastic restoring force, kx(t). In this case, 1I represents the damping frictional drag of the endolymph fluid, k is the spring constant of the cupular restoring force, m is the total mass of the otoconia, and x(t) is the linear displacement of the mass with respect to time. (Wilson & Melvill-Jones, 1979).
In contrast to the dynamic angular motions required to stimulate the semicircular canals, static head positions can actually stimulate the otoliths by tilting the macular sheets with respect to the linear acceleration of gravity (or alternatively, the gravito-inertial force, GIF). This fact introduces a level of ambiguity when trying to differentiate between head tilt and linear translations, as the same otolith signal can code for either one. On Earth, other cues, such as rotational semicircular canal inputs, the surrounding visual environs, and kinesthetic feedback, are used to differentiate between these two sensations. In a microgravity environment, however, this tilt-translation ambiguity is no longer mediated by gravitational inputs and a conflict arises between the vestibular sensations and other sensory inputs (Young & Sinha, 1998). Because of the lack of a tonic downward acceleration, head tilts no longer cause displacement of the otolith macula; only linear accelerations provide otolith inputs. Therefore, the brain learns to interpret all linear acceleration signals as translational. Upon return to the 1-g environment of Earth, the reinterpretation persists for a time, causing astronauts to feel as if they are translating across a
Erik L. rown S.M Theis 2
June 2002
room, when in fact all they are doing is tilting their head with respect to the now-present gravity (Young, et al, 1984; Parker, et al, 1985).
Visual-Vestibular Interaction
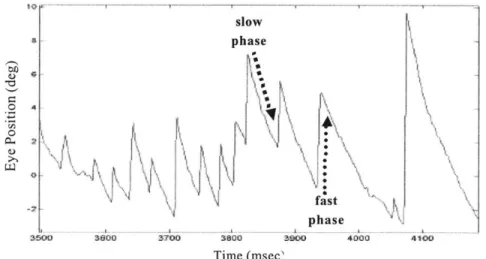
While the semicircular canals and otolith organs provide critical inputs to the body's orientation and posture control systems, they do not work alone. Visual inputs provide an important reference for body alignment, mediation cf the tilt-translation ambiguity, and target acquisition for guiding motions. The vestibulo-ocular reflex (VOR) ties together motions of the head and motions of the eyes for the purpose of gaze stabilization. For instance, in order to maintain a fixed gaze on an object during a leftward head motion, the eyes must compensate by moving towards the right. Likewise, in order to maintain a fixed gaze during a downward linear acceleration, the ocular counter-rotation must be upwards. During continuous head turns or linear accelerations, these compensatory ocular reflexes may be greater than the physical limitations of the eye socket, and saccades, a fast phase resetting of the eyeball, may occur. The slow phase drift, together with this fast phase reset is known as nystagmus. (Figure 5) By means of a central
"velocity storage" mechanism, modeled as either a positive feedback loop (Robinson, 1977) or a leaky integrator (Raphan, Matsuo, & Cohen, 1977), the time constant of the slow phase of this nystagmus is prolonged to nearly three times that of the vestibular end-organ dynamics.
slow phase 0 Ilk
IN1
fast phase 36W0 3900 3700 3800 3900 4000 4100 Time (msec,Figure 5: The fast and slow phases of a down-beating nystagmus
The fundamental open-loop reflex of VOR is one of the simplest and most primitive in the human body, tracing out a short reflex arc from the afferent pathways of the vestibular organs to the vestibular nuclei, and onward, through a short interneuron, directly to the motoneurons that drive ocular motion. With a latency of just 16-msec, pure vestibular signals between 0.5 and 5.0-Hz
June 2002
can drive compensatory eye movements with great accuracy (Leigh, 1996). Complexity is introduced, however, by other cognitive and sensory inputs, which also synapse on the wstibular nuclei and cause modifications to the overall ocular response.
Vision offers a particularly important input to this system, as moving visual scenes also provoke analogous compensatory eye motions for the purposes of stabilizing images on the retina. Large coherent motions in the visual field, such as the motion seen out the window of a train, trigger a reflexive optokinetic nystagmus (OKN) with the same characteristic sawtooth "beating" pattern as VOR. In a research setting, this motion stimulus is generally provided by immersive virtual reality environments or large optokinetic drums with alternating black and white stripes. Paige (1983a) showed a decline in OKN gain in the squirrel monkey for stimuli above 0.01-Hz. This observation provides good evidence that the evolution of OKN was at least partially driven to complement the VOR response, which falls off at low frequencies as the semicircular canals fail to transmit accurate information about rotational velocity. OKN is also subject to the same velocity storage mechanisms that prolong the time constant of VOR. This is evidenced by optokinetic afternystagmus, or OKAN, where OKN beating persists for a few seconds following removal of the optokinetic stimulus.
Fortunately for vestibular researchers, the reflexes governing nystagmus offer fairly accurate insight into the dynamic state of neurovestibular processing. On the basis of the torsion-pendulum model of the semicircular canals, the fluid dynamics of the inner ear can be deduced for a given head motion. Then, by tracking eyeball motion with induction coils, electrooculography (EOG), or a video imaging system, and comparing this ocular data with the expected vestibular output, investigators can obtain a non-invasive measure of the intervening central processes.
VOR Gain Plasticity
VOR and OKN gain can be described as efficiency metrics for gaze stabilization, measuring the ratio of compensatory eye velocity to input head or visual field velocity. Thus, a head turn matched by an equal and opposite eye rotation would be considered to have a gain of 1.0, and gaze would be perfectly stabilized. While human values for vertical and horizontal VOR gain have been calculated around 0.9-1.1 during walking, the gain of torsional VOR never even approaches 1.0, generally providing less than 65% counter-rotation. This discrepancy may be
June 2002
explained by both physical limitations and by the very root of the reflex. Both horizontal and vertical movements of the head or visual field cause displacement of visual images on the retina. This "retinal slip" in turn causes blurring and perceptual difficulties. In the case of motion about the roll axis, no linear slip is present in the fovea, and visual acuity remains high (Leigh, 1996).
Modifications to VOR gain can take two forms: transient adjustment and persistent plasticity. Transient adjustments to VOR are generally the result of temporary higher order inputs to the vestibular nuclei. Fixation on imaginary stable visual targets, for instance, has been shown to decrease gain (Barr, Schultheis, & Robinson, 1976), as have fatigue and depressed alertness
(Collins & Guedry, 1962).
Because VOR is a short-arc open-loop system with no feedback, however, mechanisms must exist to adjust this gain in response to longer-lasting changes, particularly in the cases of neurovestibular trauma, cell death due to aging, or atypical environmental conditions, such as the constant rotation onboard a centrifuge. This ability of the system to respond to changing needs and to retain that adjustment is known as plasticity (Melvill Jones, 1977; Miles & Lisberger,
1981). Vision-reversing prisms, telescopic lenses, optokinetic drums, and cross-axis training with
combined vertical and horizontal stimuli have been shown to induce plastic changes in VOR gain (Quinn, 1998). In one of the classic demonstrations of this phenomenon, Melvill Jones (1977) showed that VOR gain was attenuated with an 8-hour exposure to vision-reversing glasses, but that longer durations (approaching 4 weeks) introduced a phase change of over 160', nearly reversing the response to properly compensate for the stimulus of the glasses.
For both transient adjustments and longer-lasting plasticity, changes that serve a functional purpose (e.g.-improving vsual acuity in response to environmental or physiological demands) are termed "adaptive." Simple decreases of gain in response to repeated or prolonged stimuli are better described as "habituation."
The primary inputs for adaptive VOR gain control are fpesumed to be signals from the visual system. Persistent retinal slip indicates that there is a discrepancy in the compensatory ocular response and that a change to the gain of the system is required. Slip in the direction of head rotation reflects an over-rotation of the eyes, and points towards the need for a decrease in gain. Retinal slip counter to the direction of rotation, on the other hand, is due to under-compensation, and requires an increase in gain. Thus, visua-vestibular interaction (VVI) says a key role in
June 2002
VOR plasticity. In fact, following unilateral labyrinthectomy, "normal VVI" is necessitated for
recovery of midband VOR gain and postural stability (Paige, 1983b).
Modeling Multi-Sensory Interaction
In reality, the ocular drive known as VOR is actually a composite response of many reflexes. Interactions between the semicircular canals, otolith organs, visual system, kinesthetic apparatus, and top-down cognitive inputs combine to produce a single estimate of head velocity relative to the world. Of these interactions, canal-otolith and canal-visual are the most salient and the most commonly investigated.
Interactions between the two sets of vestibular organs are evidenced in many ways. In 1965, Guedry demonstrated that although the restoring action of the cupula seems to be gravity-independent, there is an influence of the otolith organs during constant velocity rotation about an Earth-horizontal axis. Young and Henn (1975) further explored this paradigm, rotating rhesus monkeys about varying off-vertical axes at varying angular velocities. Here, the utricle and saccule were both in constant rotation with respect to the gravito-inertial force vector, and there was a tendency towards prolonged nystagmus with a compensatory bias component. The peak-to-peak modulation of this nystagmus varied sinusoidally with increasing tilt angle, pointing towards otolith mediation.
Another interesting interaction is evidenced by the post-rotatory phenomenon of "dumping." For subjects rotated about an Earth vertical axis, a quick post-rotatory head tilt which changes the saccular alignment with respect to the gravity vector, will cause a rapid shortening of the VOR time constant (Benson & Bodin, 1966; Raphan, et al, 1981). Furman and Schor (2001) established that the velocity storage mechanisms that prolong VOR time constants are less effective in constant-velocity off-vertical axis rotation (OVAR) than in Earth-vertical axis rotation (EVAR), but more effective in sinusoidal OVAR stimulation. They hypothesized that in addition to semicircular canal inputs, the otoliths also contribute to estimates of angular head velocity by analysis of the swinging GIF vector. Low pass filtering of this estimate creates the horizontal bias described by Guedry, and reduction of the velocity storage feedback gain accounts for the mechanisms of dumping described by Raphan, et al. Interaction of these two components results in emergent properties as described above, such that otolith influences during sinusoidal rotation more than compensate for the reduced effectiveness of velocity storage.
Erika L. Brown, S.M. Thesis
29
June 2002
Modeling efforts, including those by Robinson (1977) and Paige (1983a), have pointed to a linear combination of the visual and vestibular inputs, such that for a subject rotating in a earth-fixed frame receiving veridical visual information about her rotation, this "True Visual VOR"
(TVVOR) is given as:
TVVOR
=VOR +
OKN(1-VOR)
(8)And for a subject rotating in a rotating frame, such that the world appears stable, the "Visually
Suppressed VOR" (VSVOR) is given as:
VSVOR
=VOR(1-OK)
(9)Zupan, Merfeld, and Darlot (2002) have recently extended these modeling efforts to embrace three-dimensional canal-otolith and canal-vestibular interactions as inputs to spatial orientation and eye movements. Recognizing that each sensory modality is most sensitive within a certain frequency band, this model implements a central nervous system (CNS) estimate of sensory dynamics to produce a more accurate broadband "sensory estimate" of a given variable. A second internal model of body dynamics is utilized to process these various sensory estimates of a single variable into "intermediate estimates" with a common representation, i.e. - all rotation variables in terms of angular velocity, rather than velocity and position. The third stage of the model takes a weighted sum of all the intermediate estimates of the variable in question to produce an updated CNS "central estimate" for driving slow-phase eye movements and future iterations of the feedback control loops described above. In this model, static otolith inputs are used to provide an estimate of gravity, while phasic afferents signal linear accelerations. This distinction provides the CNS with a unique separation of the two variables, rather than the usual combined gravito-inertial force vector provided by the otoliths. In addition, the changing estimates of gravity in off-axis rotation provide input to the estimate of angular velocity as discussed earlier.
The lack of visual inputs is represented, not by zero visual input, but rather by deactivating the visual feedback and smooth pursuit systems by opening those feedback pathways. This distinction is crucial in the calculations that drive adaptive VOR gain because a null input for retinal slip would indicate perfect gaze stabilization. This signal would compete with incoming