PGC-1α régule un programme onco-métabolique capable de réprimer l’agressivité du cancer de la prostate
Texte intégral
Figure





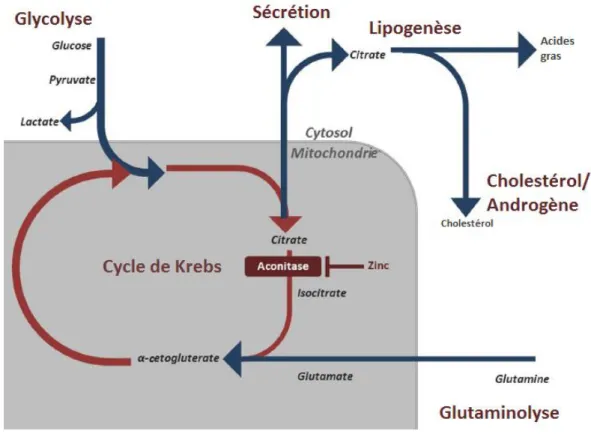

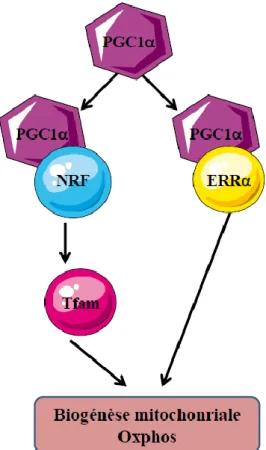

Documents relatifs
Q37 Preferred treatment in addition to ADT in patients with newly diagnosed low-volume metastatic (M1) castration-sensitive/naïve prostate cancer relapsing after local treatment of
• The number of new prostate cancer cases and deaths will almost double by 2030 in Latin America and the Caribbean if current trends continue.. Proportion of new cases
Previous work has demonstrated that membrane lipid rafts associated with key signalling proteins mediate growth and survival signalling pathways in prostate cancer
With the effectiveness of inhibitors against the Pak2 activators Rac1 and CDC42 showing an inhibition of TGF-β suggesting Pak2 activity in PC-3 cells, in order to test the
Because SgII expression is up-regulated both in vivo in high grade PCa and in vitro in androgen-deprived LNCaP cells, we next questioned whether SgII might be an essential effector
The neuropep- tide 26RFa is expressed in human prostate cancer and stimulates the neuroendocrine differentiation and the migration of androgeno-independent prostate cancer cells...
Le questionnaire, conc ¸u par le Cams, a été construit afin de recueillir de fac ¸on rétrospective les caracté- ristiques générales de la population étudiée, l’état de
The effects of cyclopamine on androgen regulated genes was common to several types of human prostate cancer cell lines grown under androgen deprived conditions so the effect was
