Fluid and particle passage in three duiker species
Texte intégral
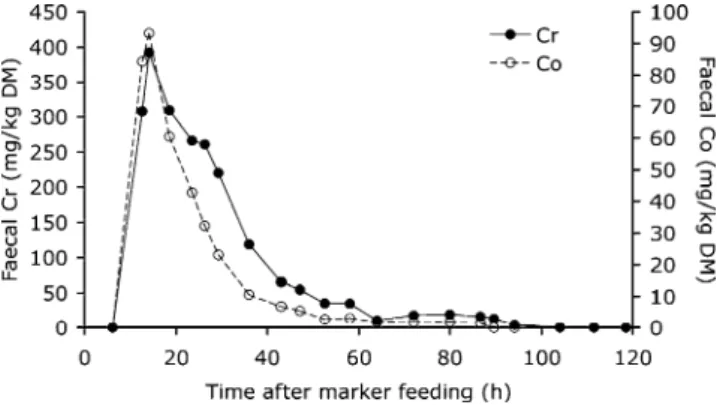
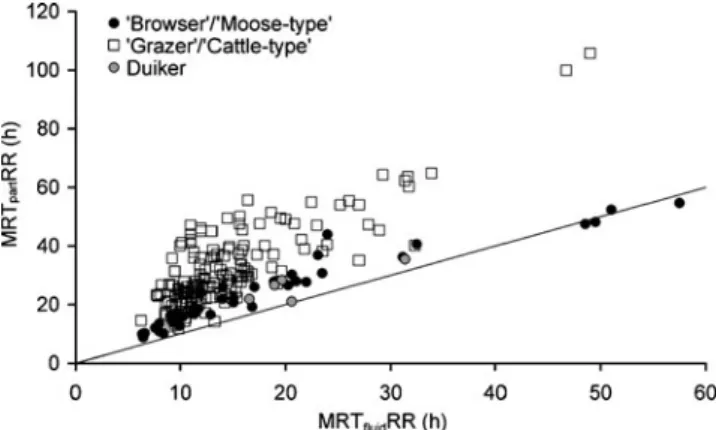
Figure
Documents relatifs
the selection aiming at a larger daily gain, which is advantageous for the feed conversion of the individual, may lead to the development of a cow stock of a
AFLP: Amplified fragment length polymorphism; CDS: Coding sequence; GO: Gene ontolology; Hrp: Hypersensitive response and pathogenicity; MLSA: Multilocus sequence analysis;
rigida (Viv.) Rothm. fontanesii Rothm., the latter two absent from the Italian territory. spinosa he distinguished three varieties based upon differ- ences in bracteole
In monogastric species the main effects of yeast supplementation are stimulation of brush border disaccharidases, anti-adhesive effect against pathogens, stimulation
(2011) explore a particularly challenging issue con- nected to HHS: they attempt to trace the colonization and recombination history of an ancient (several MYA) hybrid species,
TtsI of NGR234 and other rhizobia has been shown to control the expression of several genes containing a TB in the promoter region, but direct contact had never been shown.. To
Title: spa type distribution in Staphylococcus aureus originating from pigs, cattle and poultry..
sdiA Enterobacter cloacae Front Cell Infect Microbiol. Citrobacter
