High-cell-density phorbol ester and retinoic acid upregulate involucrin and downregulate suprabasal keratin 10 in autocrine cultures of human epidermal keratinocytes
7
0
0
Texte intégral
Figure
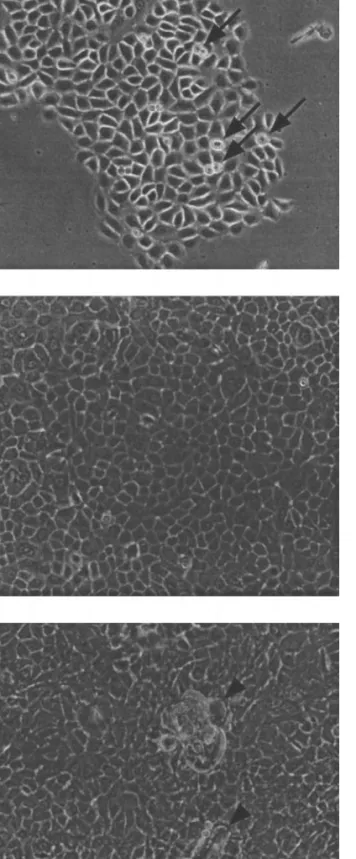
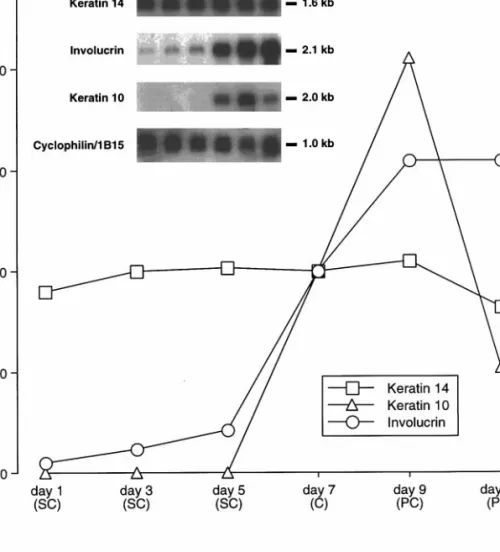
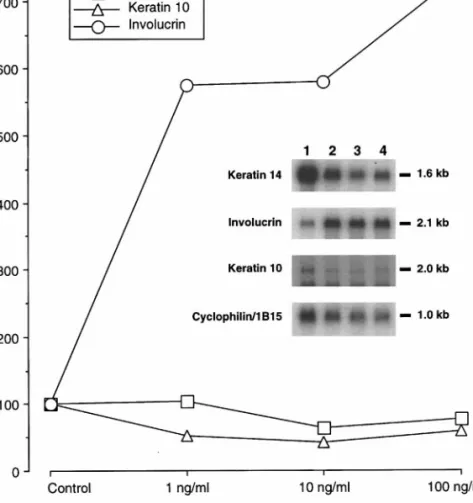
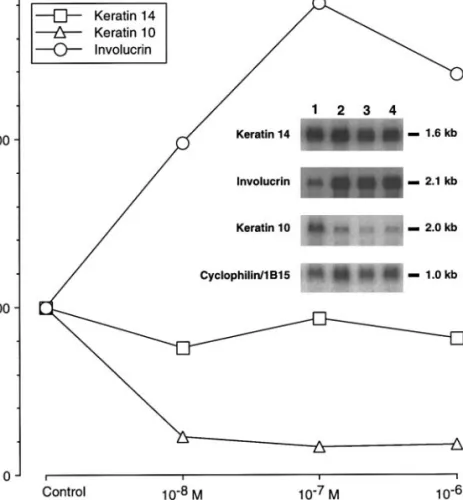
+2
Documents relatifs
