HAL Id: hal-02594479
https://hal.inrae.fr/hal-02594479
Submitted on 15 May 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Variations temporelles des peuplements de poissons des
cours d’eau français
E. Araujo Calçada
To cite this version:
E. Araujo Calçada. Variations temporelles des peuplements de poissons des cours d’eau français. Sciences de l’environnement. 2008. �hal-02594479�
RAPPORT DE STAGE DE MASTER 1
ECOLOGIE ET ENVIRONNEMENT
par Emmanuelle Araujo Calçada
Sujet :
Variations temporelles des peuplements de poissons
des cours d'eau français.
Maître de stage : Jérôme Belliard
Enseignant responsable : Jean-Christophe Lata
UNITE HYDROSYSTEMES ET BIOPROCEDES
EQUIPE HYDRECO
Cemagref - Unité HBAN Parc de Tourvoie, BP44 92 163 Antony Année 2008 CemOA : archive ouverte d'Irstea / Cemagref
REMERCIEMENTS CemOA : archive ouverte d'Irstea / Cemagref
SIGLES et ABBREVIATIONS
Cemagref : CEntre national du Machinisme Agricole, du Génie Rural, des Eaux et des Forêts
HBAN : Hydrosystèmes et Bioprocédés d'Antony
HYDRECO : HYDRoECOlogie des cours d'eau
DCE : Directive Cadre européenne sur l'Eau
RHP : Réseau Hydrobiologique et Piscicole
CSP : Conseil Supérieur de la Pêche
ONEMA : Office National de l'Eau et des Milieux Aquatiques
HER : hydro-écorégion
CIEM : Conseil International pour l'Exploration de la Mer
CemOA
: archive
ouverte
d'Irstea
RESUME / ABSTRACT
Ce stage porte sur la variabilité temporelle des peuplements de poissons des cours d'eau français appréhendée à travers l'étude d'un important jeu de données. Les analyses sont effectuées sur des données issues de 473 stations réparties sur le territoire français échantillonnées annuellement sur une période de neuf ans (de 1995 à 2003). Elles se basent sur des méthodes déjà employées pour des problématiques similaires, à savoir l'étude des coefficients de variation et des turn-over. A travers ces analyses, la variabilité des peuplements est évaluée et reliée à différents facteurs potentiellement explicatifs qui sont de trois types : les caractéristiques environnementales des stations, les perturbations anthropiques et l'échantillonnage. Puis, la variabilité des populations est étudiée en fonction des caractéristiques biologiques et écologiques des espèces et par rapport à la position de l'espèce vis-vis de son optimum écologique. Il ressort de ces analyses que la variabilité des peuplements de poissons échantillonnés par pêche électrique dépend à la fois de l'environnement naturel, des altérations d'origine anthropique (qualité de l'habitat) et de l'échantillonnage. Il ressort également que les espèces de grande taille, à fortes fécondité et longévité ont des populations plus stables. Enfin, il semble que, pour certaines espèces, les populations sont d'autant plus stables qu'elles sont proches de leur optimum écologique.
This internship deals with temporal variability in stream-fish assemblages in France. Data concern 475 sites sampled annually over a nine-year period from 1995 to 2003. By calculating coefficients of variation and turn-over, this study uses already known methods. Through these analyses, we assess temporal variability of assemblages and then link this variability to three potential explanatory factors: environmental site attributes, anthropogenic disturbances and sampling factors. Then, variability, studied at the population scale, is related to biological and ecological characteristics of species and to the location of the specie in relation to its ecological optimum. These analyses indicate that fish assemblage variability depends on natural environment, anthropogenic modifications (quality of habitat) and sampling factors. Moreover, the results show that large species, with high fecundity and longevity, have the most stable populations. Finally, for several species, populations near of their ecological optimum seem to be the most stable.
CemOA
: archive
ouverte
d'Irstea
TABLE DES MATIERES
INTRODUCTION ... 6
- Présentation de l'organisme d'accueil ... 6
- Présentation du sujet et de son contexte ... 8
MATERIEL ET METHODES... 10
Présentation des données ... 10
- Les données poissons... 11
- Les évaluateurs de la variabilité des peuplements : les CV et les turn-over ... 11
- Les données sur la nature de l'intensité d'échantillonnage... 13
- Les données environnementales... 13
- Les caractéristiques biologiques des espèces ... 16
- Les probabilités de présence des espèces à l'échelle stationnelle ... 17
Méthodes d'analyse... 18
RESULTATS ET DISCUSSION ... 19
Evolution de la fréquence des espèces au cours du temps ... 19
Analyse de la variabilité des peuplements et des différences entre stations ... 22
- Quantification des variabilités : CV et turn-over ... 22
- Influence de la technique d'échantillonnage sur la variabilité des peuplements ... 25
- Variabilité des peuplements en fonction des caractéristiques des stations et de l'échantillonnage... 26
Analyse de la variabilité des populations et des différences entre espèces... 31
- Variations du CVDE selon les caractéristiques des espèces... 31
- Variabilité d'une espèce selon sa position vis-à-vis de son optimum écologique.... 33
CONCLUSION ... 35
BIBLIOGRAPHIE ... 36
ANNEXES ... 36
CemOA : archive ouverte d'Irstea / CemagrefINTRODUCTION
Présentation de l'organisme d'accueil
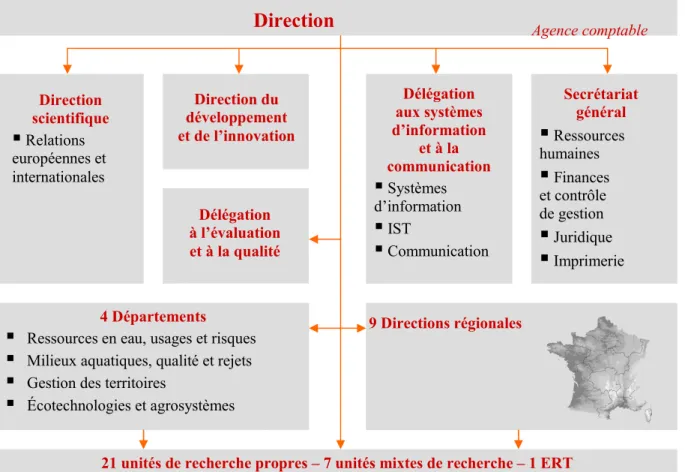
Le Cemagref, créé en 1981, est un établissement public à caractère scientifique et technique (EPST) sous la double tutelle des ministères en charge de la Recherche et de l’Agriculture. C’est un organisme de recherche finalisée sur la gestion des eaux et des territoires. Il centre ses recherches sur les ressources en eau de surface, les systèmes écologiques aquatiques et terrestres, les espaces à dominante rurale, les technologies pour l’eau, les agro-systèmes et la sûreté des aliments. Il crée des méthodes et des innovations technologiques pour l’aide à la décision et la gestion de systèmes naturels et anthropisés. La compréhension et la caractérisation des systèmes étudiés s’appuient fortement sur l’expérimentation et la modélisation. Ses départements et laboratoires sont définis par un objet de recherche et non par une discipline (fig.1).
Figure 1 : Organigramme du Cemagref
4 Départements
Ressources en eau, usages et risques Milieux aquatiques, qualité et rejets Gestion des territoires
Écotechnologies et agrosystèmes
Direction
générale
Direction scientifique Relations européennes et internationales Direction du développement et de l’innovation Délégation à l’évaluation et à la qualité Délégation aux systèmes d’information et à la communication Systèmes d’information IST Communication Secrétariat général Ressources humaines Finances et contrôle de gestion Juridique Imprimerie Agence comptable 9 Directions régionales21 unités de recherche propres – 7 unités mixtes de recherche – 1 ERT
CemOA
: archive
ouverte
d'Irstea
L'unité de recherche Hydrosystèmes et Bioprocédés du groupement d'ANtony, HBAN, constitue le pôle Eau du centre d'Antony. Elle appartient au département "Ressources en eau, usages et risques". Les travaux de recherche de cette unité sont appliqués à des domaines aux forts enjeux environnementaux. Les recherches portent sur le fonctionnement hydrologique (relations bassins versants - débits), sur l’impact des aménagements hydro-agricoles et des pratiques agricoles (depuis la parcelle jusqu'au bassin versant aménagé), sur le fonctionnement des écosystèmes fluviaux (peuplements piscicoles en particulier), sur les processus de dégradation et de transformation (eaux usées en stations d'épuration, eaux naturelles) ainsi que sur l’impact éco-toxicologique de ces rejets. Elles débouchent sur une expertise globale sur le continuum "pluie, réseaux hydrographiques-systèmes d'assainissement, débits et qualité des cours d'eau" pour une gestion intégrée des bassins versants et des hydrosystèmes.
L’unité développe également des recherches sur le fonctionnement des installations de stockage de déchets, du point de vue des transferts d'eau et de lixiviats (dans les systèmes de couverture, de drainage-recirculation et d'étanchéité), de l'influence de ces transferts sur l'évolution des déchets, ainsi que de la gestion durable du risque environnemental associé. Les modèles et l'expertise de l'unité s'appliquent à l'ingénierie des stations d'épuration et des centres de stockage des déchets, à la prévision des ressources en eau (fleuves, retenues) et des événements exceptionnels (crues, étiages), à la gestion des populations de poissons et de l'occupation des bassins versants fortement anthropisés et à la biogéochimie des cours d’eau (polluants d’origine agricole, contaminants organiques et métalliques).
L’équipe Hydroécologie des cours d’eau (HYDRECO), qui m'accueille au sein de l'unité HBAN, a pour thème de recherche les déterminants physiques du fonctionnement écologique des hydrosystèmes d’eau courante. Le but est l’étude du fonctionnement des écosystèmes aquatiques, appréhendé à travers les peuplements de poissons. L’objectif est d'évaluer les effets des activités humaines sur ce fonctionnement. A terme, l'ambition serait de développer des outils d'aide à la décision et à l'évaluation des choix technologiques en matière d'aménagement et de réhabilitation, en vue d'une gestion équilibrée du patrimoine aquatique. Les travaux de l'équipe portent essentiellement sur le déterminisme des relations entre les poissons et leur habitat. Ces relations sont en effet indispensables pour mieux connaître et comprendre l'influence des différents facteurs qui régissent les processus d'organisation et de structuration des peuplements piscicoles, afin de pouvoir prévoir les conséquences des
CemOA
: archive
ouverte
d'Irstea
modifications d'origine naturelle ou anthropique, auxquelles sont soumis les milieux aquatiques continentaux.
Présentation du sujet et de son contexte
La Directive cadre européenne sur l'eau (DCE), récemment mise en place, a placé le fonctionnement écologique des hydrosystèmes au centre des préoccupations en matière de politique de l'eau. L'objectif pour la France est le bon état des eaux (cours d'eau, plans d'eau, eaux de transition et eaux côtières) d'ici 2015. Cette directive se traduit donc par la nécessité d'une évaluation de l'état écologique des eaux. Cette évaluation comprend d'une part des paramètres physico-chimiques et d'autre part des paramètres biologiques (bioindicateurs). Concernant les cours d'eau, les peuplements de poissons sont identifiés comme l'un des éléments biologiques clés autour desquels devront s'organiser la surveillance et la gestion des milieux. En effet, les poissons dépendent étroitement de multiples paramètres de leur milieu écologique qui s'expriment à différentes échelles de temps et d'espace. Leurs peuplements traduisent notamment la diversité des habitats offerts et le fonctionnement hydrologique des cours d'eau. Ils peuvent ainsi être considérés comme de bons intégrateurs du fonctionnement global des hydrosystèmes fluviaux dont ils constituent une bonne expression de l'état de santé. Atteindre les objectifs fixés par la DCE nécessite donc de connaitre précisément les facteurs qui contrôlent la structure et la dynamique des peuplements de poissons. L'évaluation de l'état des cours d'eau sur la base de la composition des peuplements en place permet ensuite de proposer des mesures de gestion adaptées assurant leur conservation ou leur restauration. En outre, les connaissances dans ce domaine via la variabilité temporelle sont encore peu développées et cette étude a également toute sa place dans une optique de recherche fondamentale.
En effet, l'organisation spatiale des peuplements de poissons à l'intérieur des réseaux hydrographiques et les facteurs naturels et anthropiques qui les contrôlent ont fait l'objet d'une littérature scientifique abondante (Matthews, 1998). En revanche, les aspects liés à leurs variations temporelles ont été beaucoup moins abordés en raison, notamment, d'un manque de données pertinentes (suivis à long terme) pour aborder ces questions.
Ce stage consiste à évaluer la variabilité temporelle des peuplements de poissons des cours d'eau français en détaillant plus particulièrement l'évaluation des phénomènes d'extinction et de colonisation. Cependant ces derniers aspects n'ont pas encore été abordés en détail pour le
CemOA
: archive
ouverte
d'Irstea
d'un jeu de données acquis dans le cadre du Réseau Hydrobiologique et Piscicole (RHP) mené par le Conseil Supérieur de la Pêche (CSP, nouvellement ONEMA) qui regroupe plus de 700 stations dont les peuplements de poissons ont fait l'objet d'un suivi annuel sur une période d'une dizaine d'année. Ce jeu de données, par son ampleur, permet donc d'aborder les phénomènes temporels sur une large échelle spatiale et pour une grande diversité de cours d'eau.
A ce stade, mon travail de stage a donné lieu à un grand nombre d'analyses et de traitement statistiques. Seuls les résultats les plus significatifs et pertinents sont présentés dans le cadre du rapport.
Les objectifs de l'étude peuvent être posés sous la forme des quatre points suivants :
1. Caractériser et quantifier la variabilité des peuplements et des populations
2. Evaluer dans quelle mesure la variabilité des peuplements dépend (1) des caractéristiques naturelles du cours d'eau et de son environnement (sa taille notamment); (2) de la nature et de l'intensité des modifications anthropiques (qualité d'eau et d'habitat); (3) d'aspects liés à l'échantillonnage
3. Examiner la variabilité des populations en fonction des caractéristiques biologiques et écologiques des espèces
4. Pour une espèce donnée, vérifier dans quelle mesure la variabilité temporelle de ses populations varie selon sa situation plus ou moins centrale vis-à-vis de son optimum écologique CemOA : archive ouverte d'Irstea / Cemagref
MATERIEL ET METHODES
Présentation des données
Le jeu de données sur lequel se base ce stage a été obtenu par le Réseau Hydrobiologique et Piscicole (RHP) du Conseil Supérieur de la Pêche. Il concerne l'état annuel des peuplements de poissons des cours d'eau français. Les données ont été acquises sur plus de 700 stations de 1995 à 2003 et couvrent donc 9 années. Chaque station correspond à un tronçon défini de cours d'eau dont la taille dépend de la largeur du cours d'eau. Sur chaque station, l'échantillonnage du peuplement est réalisé par pêche électrique. Selon la largeur et la profondeur du cours d'eau, deux techniques différentes sont appliquées : lorsque les conditions le permettent (petits cours d'eau et milieux peu profonds), une prospection complète de la station, à pied, est retenue (plusieurs passages successifs peuvent alors être réalisés afin de disposer d'une estimation de l'effectif exact de poissons présents sur la station); en revanche, pour les grands cours d'eau trop profonds pour être échantillonnés à pied, la pêche en bateau partielle (le long des rives) est privilégiée. Le type d'échantillonnage pratiqué sur les petits cours d'eau, donne une représentation assez précise du peuplement en place (OBERDORFF et al., 2002, d'après PALLER, 1995; ANGERMEIER et SMOGOR, 1995). S'agissant des grands cours d'eau, la pêche en bateau partielle le long des rives donne une bonne représentation de la richesse en espèces de la station mais une image imparfaite et biaisée de la structure du peuplement en place (OBERDORFF et al., 2002, d'après CHANCEREL, 1992). Cet aspect devra être pris en compte dans l'interprétation des résultats. Pour les pêches à plusieurs passages, seuls les résultats du premier passage ont été conservés afin d'assurer une plus grande cohérence de l'ensemble du jeu de données.
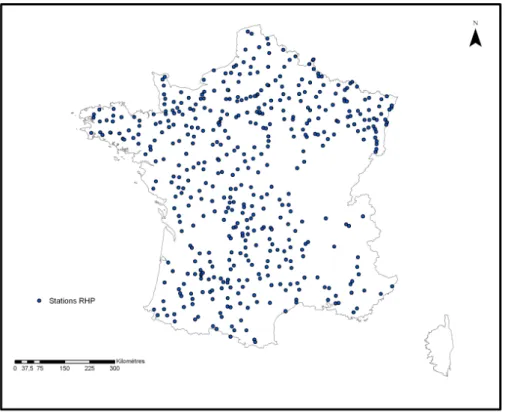
Initialement, les mesures sur le terrain ont été coordonnées au niveau régional par le CSP. Le but était d'obtenir une bonne représentativité de la réalité des cours d'eau français à travers l'échantillonnage. Les stations sont donc, en théorie, réparties de façon homogène dans chaque région et dans les types de cours d'eau (plus ou moins aménagés, le long du gradient longitudinal…). Toutefois, seules les stations comportant des données pour les 9 années étudiées (de 1995 à 2003) ont été conservées. De même, les stations situées sur les milieux artificiels de type canaux ont été éliminées. Au final 473 stations ont été sélectionnées, chacune disposant de 9 années de suivi. Ce nombre de stations reste considérable et permet une étude à grande échelle. La distribution des stations retenues sur le territoire français est
CemOA
: archive
ouverte
d'Irstea
présentée sur la carte suivante (fig.2). Nous constatons que leur distribution est globalement homogène à l'exception d'une large partie présentant une densité de stations très faible et de la Corse pour laquelle la procédure de sélection des données n'a permis de retenir aucune station RHP.
Figure 2 : Carte des stations étudiées
Les espèces (ou groupes d'espèces) étudiées sont au nombre de 57, elles sont recensées en Annexe 1. Certaines espèces ont été regroupées en raison de problèmes de détermination sur le terrain : il s'agit des lamproies du genre lampetra (lamproie de planer et lamproie de rivière) et des deux phénotypes de truites (truite commune et truite de mer).
- Les données poissons
Elles sont utilisées de deux manières : soit en terme d'indice de densité (nombre d'individus capturés par mètre carré), soit en présence/absence sans prendre en compte le nombre d'individus (présence=1, absence=0).
- Les évaluateurs de la variabilité des peuplements : les CV et les turn-over
Afin d'analyser la variabilité temporelle des peuplements de poissons, nous nous intéressons dans un premier temps à leur stabilité, c'est-à-dire au maintien des densités des espèces au cours du temps. Pour cela, nous calculons différents types de coefficients de variation (CV).
CemOA
: archive
ouverte
d'Irstea
Le CV est donné par la formule : ×100 − = moyenne type écart CV
Cinq types de coefficients de variation sont calculés :
Le CVRSL qui est un CV calculé à partir des richesses spécifiques pour chaque station (locales). Il nous donne une idée de la stabilité dans le temps du nombre d'espèces pour une station donnée.
Le CVDT qui est un CV calculé à partir de la densité totale du peuplement pour chaque station. Il nous donne une idée de la stabilité dans le temps de la densité des peuplements, sans distinction d'espèces, pour une station donnée.
Le CVSP qui est un CV de densité de chaque espèce. Pour chaque station, la densité de chaque espèce est calculée pour chaque année (nombre d'individus / surface échantillonnée) puis un CVSP est calculé pour chaque espèce à partir des densités.
Le CVDM qui est un CV moyen calculé pour chaque station à partir des CVSP. Pour cela, nous choisissons de ne considérer que les espèces présentes au moins 50% de la période analysée afin de limiter l'effet des espèces rares dans le calcul de la variabilité du peuplement (HUGUENY et al., 1995). Le CVDM nous donne une idée de la stabilité moyenne des espèces dans le temps pour une station donnée.
Le CVDE qui est un CV moyen par espèce. Il est utilisé pour l'étude des variations en fonction des caractéristiques des espèces. Les analyses ne portent que sur les cas où l'espèce est présente les neuf années consécutives dans une station. Un coefficient de variation (CVSP) est recalculé par espèce et par station (quand l'espèce est présente neuf ans) puis la moyenne de ces CV donne un CVDE par espèce qui va être le paramètre utilisé dans les analyses. Le principe est donc identique au calcul du CVDM précédent, sauf que cette fois-ci l'espèce doit être présente durant toute la période étudiée et que la moyenne ne se fait plus par station mais par espèce.
Un schéma illustrant les différentes étapes des calculs de CV est fourni en Annexe 2.
CemOA
: archive
ouverte
d'Irstea
Dans un second temps et en complément des indicateurs précédents, nous nous intéressons au renouvellement des peuplements afin de savoir si nous pouvons observer ou non un maintien des espèces au cours du temps. Cela est mesuré à partir de l'index de turn-over (renouvellement) faunistique (BELKESSAM, 1998, d'après DIAMOND et MAY, 1977; MEFFE et BERRA, 1988). Le turn-over entre deux échantillons est mesuré à partir du
rapport:
(
)
(
S1 S2)
E C T + += , C et E étant respectivement le nombre de colonisation et d'extinction des espèces durant la période d'étude, S1 et S2 le nombre d'espèces de chaque échantillon. T varie de 0, pas de turn-over, à 1, turn-over total. A partir des données concernant la présence ou l'absence des espèces, un turn-over est calculé pour chaque année à chaque station. Pour la majorité des analyses un turn-over moyen est calculé par station (T moyen).
- Les données sur la nature et l'intensité de l'échantillonnage
Dans le but de disposer d'une variable quantitative qui prenne en compte l'effet de l'échantillonnage, le paramètre suivant est calculé à partir des données :
• le ratio échantillonné = surface échantillonnée / surface totale de la station Nous disposons en outre d'un paramètre sur la méthode d'échantillonnage :
• le type de prospection (à pied ou en bateau)
- Les données environnementales
Huit variables quantitatives permettant de décrire les caractéristiques naturelles de cours d'eau ont été évaluées sur chaque station :
• la surface du bassin versant exprimée en km2 • la distance à la source exprimée en km • la largeur du cours d'eau exprimée en m • la pente de la station (IGN) exprimée en ‰ • la profondeur moyenne exprimée en m • l'altitude exprimée en m
• la température moyenne de janvier exprimée en °C (Tjanvier) • la température moyenne de juillet exprimée en °C (Tjuillet)
CemOA
: archive
ouverte
d'Irstea
Afin de s'affranchir des liens de dépendance entre les paramètres, cinq variables composites décrivant les stations sont construites à partir des huit variables précédentes en se basant sur la méthode d'élaboration de l'Indice Poisson Rivière (IPR) (OBERDORFF et al., 2002) :
• un paramètre décrivant la position de la station dans le gradient longitudinal : G = 3,015 - 0,347 x log(Surface Bassin Versant) - 0,543 x log(Distance à la Source) Nous pouvons noter que ce paramètre varie en sens inverse de la taille du cours d'eau.
• un indice de vitesse des écoulements :
V = log(Largeur) + log(Profondeur) + log(Pente) - log(Largeur + 2 x Profondeur)
• un paramètre lié à l’altitude de la station : A = log(Altitude)
• un paramètre lié à la température moyenne de l’air : T1 = Tjuillet + Tjanvier
• un paramètre lié à l’amplitude thermique saisonnière : T2 = Tjuillet – Tjanvier
Par ailleurs, deux variables qualitatives définies sur des bases cartographiques ont également été utilisées :
• l'unité hydrologique où se trouve la station • l'hydro-écorégion (HER)
Les unités hydrologiques sont des descripteurs à l'échelle régionale. Il s'agit de bassins ou de groupes de bassins distingués sur la base de critères faunistiques. Huit ensembles ont été définis (fig. 4) : bassins mer du Nord (H1), bassin Seine (H2), bassins Manche (H3), bassins Atlantique (H4), bassin Loire (H5), bassin Garonne (H6), bassin Rhône (H7), bassins Méditerranéens (H8). CemOA : archive ouverte d'Irstea / Cemagref
Figure 4 : Carte de France donnant la position des huit unités hydrologiques
Les hydro-écorégions (fig.5) correspondent à une régionalisation des écosystèmes d'eau courante à partir des déterminants primaires géophysiques et climatiques. Le découpage par HER (WASSON, 2002) semble plus pertinent que le découpage par bassin car il tient compte de nombreux facteurs qui agissent sur les écosystèmes d'eaux courantes et donc a priori sur les peuplements de poissons. De plus, les hydro-écorégions permettent d'intégrer des effets hydrologiques et climatiques sur lesquels nous ne disposons pas de données pour le moment.
Figure 5 : HER de niveau 1
Les analyses se basent donc sur l'étude des HER. Afin d'éviter d'avoir des effectifs trop faibles dans certaines HER, les 21 HER de niveau 1 existantes sont regroupées dans 6 groupes.
CemOA
: archive
ouverte
d'Irstea
- groupe 1 : dépôts sédimentaires et alluvionnaires (Alsace, Plaine de Saône, Landes, Dépôts argilo-sableux, Dépressions sédimentaires)
- groupe 2 : plaines-collines calcaires (Coteaux aquitains, Causses, Tables calcaires, Côtes calcaires est)
- groupe 3 : formations périphériques des massifs (Jura-Préalpes du nord, Préalpes du sud, Grands Causses)
- groupe 4 : massifs anciens (Massif Central nord, Massif Central sud, Massif armoricain, Ardennes, Cévennes, Vosges)
- groupe 5 : massifs récents (Alpes internes, Pyrénées) - groupe 6 : Méditerranée
Enfin, d'autres paramètres acquis par les agents de l'ONEMA dans le cadre du programme européen EFI+ (Improvement and Spatial extension of the European Fish Index) ont également été rajoutés au jeu de données pour rendre compte de la nature et de l'ampleur des pressions anthropiques :
• la qualité de l'eau (mesurée par son état de dégradation : faible, moyen, fort) basée uniquement sur les pollutions organiques (eutrophisation…)
• la qualité de l'habitat (mesurée par son état d'altération : nul, faible, intermédiaire, fort) Finalement, les analyses incluent neufparamètres :
- G, V, A, T1 et T2
- les hydro-écorégions (HER) - le ratio échantillonné - l'état d'altération de l'habitat
- l'état de dégradation de la qualité de l'eau
- Les caractéristiques biologiques des espèces
En complément du jeu de données principal, nous disposons d'un autre jeu de données concernant les traits d'histoire de vie des espèces. Il a lui aussi été acquis dans le cadre du programme européen EFI+. Ces données ont tout d'abord été mises en forme avec des données du jeu principal. Elles ont ensuite été utilisées pour les analyses afin de faire le lien entre la variabilité des populations et la biologie et l'écologie des espèces.
CemOA
: archive
ouverte
d'Irstea
Ces données, par espèce, concernent de nombreux paramètres :
• l'habitat de vie
• l'habitat d'alimentation
• la guilde trophique de l'adulte • la guilde de migration
• la guilde reproductive • l'habitat de frai préférentiel • le comportement reproductif • les soins parentaux
• la taille maximale connue (mm) • la longévité (année)
• la fécondité absolue
• la fécondité relative (par kg de femelle) • le diamètre des œufs (mm)
• l'âge à maturité (année)
• la durée d'incubation des œufs (jour)
Un tableau recensant ces paramètres et leurs modalités est fourni en Annexe 3.
- Les probabilités de présence des espèces à l'échelle stationnelle
Grâce aux modèles développés dans le cadre de l'indice poisson rivière (IPR) (OBERDORFF et al., 2002), il est possible de calculer des probabilités d'occurrence de 34 espèces en fonction des caractéristiques naturelles des cours d'eau (ces modèles prennent en compte les 5 variables synthétiques précédemment citées G, V, A, T1, T2 ainsi que l'unité hydrologique). Par la suite, ces valeurs de probabilités seront utilisées comme des mesures à l'échelle de la station de la position de l'espèce par rapport à son optimum écologique. Une valeur de probabilité proche de 1 signifie que les conditions sur la station sont proches de l'optimum écologique de l'espèce. A l'inverse, ces conditions s'écartent d'autant plus de l'optimum que la probabilité tend vers 0.
CemOA
: archive
ouverte
d'Irstea
Méthodes d'analyse
Le logiciel utilisé pour les analyses est R. Dans l'étude des CV comme dans celle des turn-over, l'analyse est effectuée en deux étapes. Tout d'abord les tests sont réalisés facteur par facteur, à l'aide de régressions simples ou d'ANOVA, pour éclaircir le jeu de données et voir les premières tendances qui se dégagent. Ensuite les facteurs sont testés par des régressions multiples pour examiner les effets conjoints des différents facteurs et ne retenir que ceux étant réellement explicatifs. Ces régressions sont des régressions séquentielles ou régressions pas à pas, basées sur l'utilisation du critère d'information de Akaike (AIC). Le logiciel R cherche le modèle optimal expliquant la variabilité observée en gardant un bon compromis entre déviance et degrés de liberté. Pour cela il part au départ d'un modèle incluant toutes les variables et les élimine une à une jusqu'à ce qu'il n'y ait plus de diminution possible de l'AIC. Dans la procédure de sélection des variables, nous avons imposé une contrainte supplémentaire basée sur les probabilités critiques (p-value) afin de ne garder dans le modèle final que les variables qui sont vraiment explicatives (p < 0,05).
Ce type d'analyse permet de s'assurer que, si un facteur significatif dans une régression simple le reste à l'issue d'une régression multiple, cela signifie qu'il apporte réellement une explication aux observations et qu'il ne s'agit pas simplement d'un artéfact lié à des auto-corrélations entre facteurs explicatifs.
Pour les analyses par station les tests sont effectués pour les trois types de coefficients de variation (CVDM, CVDT, CVRSL) ainsi que pour un turn-over moyen calculé par station (T moyen). Pour les analyses par espèce, les tests sont effectués avec CVDE dans le cas des caractéristiques biologiques et avec CVSP dans le cas des probabilités de présence.
Compte tenu de la grande quantité de tests réalisés, seule la partie la plus pertinente des tests est présentée dans ce rapport. En outre, pour les mêmes raisons, les résultats sont présentés sous une forme synthétique.
CemOA
: archive
ouverte
d'Irstea
RESULTATS ET DISCUSSION
Evolution de la fréquence des espèces au cours du temps
Le but de ce stage n'étant pas l'étude des espèces seules mais celle des peuplements de poissons, cette partie a essentiellement un rôle introductif.
L'étude de l'évolution de la fréquence de chaque espèce au cours du temps constitue une première approche intéressante de ce jeu de données. Cela peut permettre de faire ressortir certaines tendances quant à la répartition de chaque espèce. De plus, cela permet de visualiser une partie de la variabilité des espèces au cours du temps et d'avoir une idée de la variabilité que nous pourrons observer au niveau des peuplements.
Tout d'abord, le nombre de stations occupées par une espèce par an est calculé à partir du jeu de données. Ensuite, les graphiques correspondants (nombre de stations occupées en fonction de l'année) sont tracés afin de se faire une première idée des résultats. Ces données sont alors testées statistiquement à l'aide de tests de régression pour évaluer les tendances à l'augmentation ou à la diminution qui se dégagent. Les résultats des tests sont généralement en concordance avec les déductions faites à partir des graphiques. Ils seront ensuite discutés pour chaque espèce et reliés à des facteurs potentiellement explicatifs. A ce stade, seuls certains résultats sont présentés ainsi qu'une ébauche de discussion. En effet, l'analyse approfondie de ces observations n'a pas encore été effectuée et constituera l'un des objectifs de la suite de ce stage.
Quelques graphiques particulièrement informatifs sont présentés ci-après (fig. 6 à 8). Ils illustrent à la fois des tendances à long terme sur les neuf années étudiées et des variations plus irrégulières d'une année sur l'autre.
Fréquence de l'Anguille 240 245 250 255 260 1995 1997 1999 2001 2003 Année N o m b re d e s ta ti o n s o c c u p é e s
Fréquence du Silure glane
0 5 10 15 20 25 30 35 1995 1997 1999 2001 2003 Année N o m b re d e s ta ti o n s o c c u p é e s
Figure 6 : Fréquence de l'anguille Figure 7 : Fréquence du silure glane
CemOA
: archive
ouverte
d'Irstea
Le graphique (fig.6) représente la fréquence de l'anguille au cours du temps. Nous voyons une alternance de pics et de creux semblant indiquer des variations cycliques qui illustrent surtout l'existence de variations interannuelles au-delà des évolutions à plus long terme. Toutefois globalement le nombre de stations occupées semble diminuer. Depuis les années 70, une régression des stocks a été constatée sur l'ensemble de l'aire continentale de répartition de l'anguille, qui est désormais considérée comme vulnérable par le CIEM (Conseil International pour l'Exploration de la Mer). Le graphique semble bien confirmer ce constat. Cette espèce étant un migrateur amphihalin thalassotoque, elle doit être gérée de façon particulière, au niveau européen.
Le graphique (fig.7) concerne une espèce introduite, le silure glane. Introduit au départ en Alsace, il s'est étendu récemment dans les bassins de la Seine, de la Loire et de la Garonne. Le graphique indique qu'il était assez peu présent jusqu'en 1999, date à laquelle le nombre de stations qu'il occupait a doublé. Il est ensuite en nette augmentation depuis 2000.
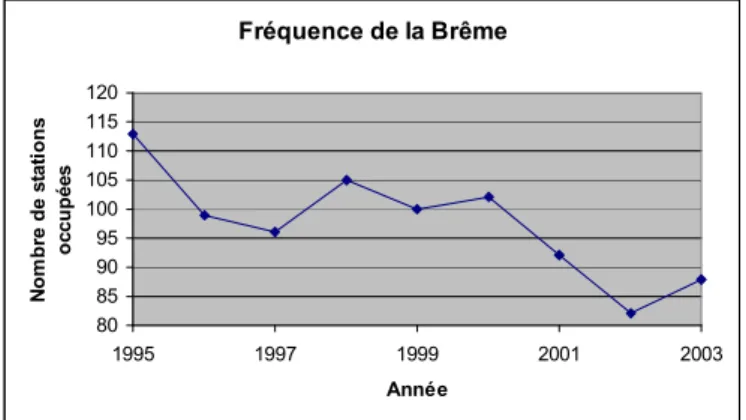
Ces résultats ainsi que ceux obtenus pour les autres espèces confirment certains constats déjà effectués et montrent également des tendances difficiles à expliquer à ce stade, comme la régression de la brème commune, espèce d'aval de cours d'eau qui supporte assez bien les pollutions (fig. 8). Fréquence de la Brême 80 85 90 95 100 105 110 115 120 1995 1997 1999 2001 2003 Année N o m b re d e s ta ti o n s o c c u p é e s
Figure 8 : Fréquence de la brème commune
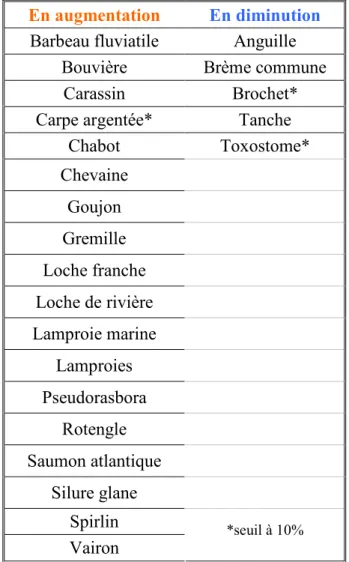
Les résultats obtenus sont présentés dans le tableau suivant (tableau 1). Les espèces n'apparaissant pas dans ce tableau sont celles pour lesquelles aucune tendance particulière n'a été montrée.
Compte tenu du faible nombre de points par espèce, un seuil de 10% a été retenu. Cependant, pour la majorité des espèces présentes dans le tableau le test est significatif au seuil 5%; les 3 espèces dont ce n'est pas le cas sont signalées par un astérisque.
CemOA
: archive
ouverte
d'Irstea
Tableau 1 : Résultats sur la fréquence des espèces
L'information principale de ce tableau est que le nombre d'espèces en régression est assez faible, puisqu'il s'agit de 5 espèces sur les 57 étudiées. Alors que le constat avait déjà été fait auparavant pour l'anguille et le brochet, il est plus étonnant de trouver ici des espèces comme la brème commune ou la tanche considérées comme tolérantes face aux altérations des cours d'eau et notamment aux pollutions.
En revanche, le nombre d'espèces en augmentation est assez important, puisqu'il s'agit de 18 espèces. Parmi celles-ci, nous retrouvons des espèces récemment introduites comme le pseudorasbora ou le silure glane, mais aussi des espèces comme la loche franche ou la bouvière pour lesquelles nous n'avons a priori pas d'explication.
A partir des résultats obtenus, il est nécessaire de récolter des informations sur chaque espèce et de croiser les données entre elles afin de tenter d'expliquer ces observations. Par exemple, il semble que plusieurs des espèces en augmentation soient des espèces rhéophiles, qui vivent plutôt en amont des cours d'eau et qui apprécient une bonne teneur en oxygène dissous et pas trop de matière organique (barbeau fluviatile, chabot, saumon atlantique, spirlin, vairon). Il pourrait ainsi s'agir d'une conséquence de l'amélioration de la qualité des eaux (notamment grâce à une diminution de phosphates). Cela pourrait aller de paire avec une diminution des espèces qui supportent bien une qualité d'eau dégradée (cas de la brème commune et de la tanche), les situations les plus dégradées s'étant globalement améliorées en cours de suivi.
En augmentation En diminution
Barbeau fluviatile Anguille
Bouvière Brème commune
Carassin Brochet*
Carpe argentée* Tanche
Chabot Toxostome* Chevaine Goujon Gremille Loche franche Loche de rivière Lamproie marine Lamproies Pseudorasbora Rotengle Saumon atlantique Silure glane Spirlin Vairon *seuil à 10% CemOA : archive ouverte d'Irstea / Cemagref
Analyse de la variabilité des peuplements et des différences entre stations
- Quantification des variabilités : CV et turn-over
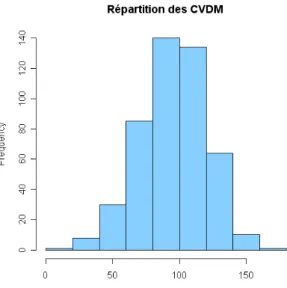
Tout d'abord, les graphiques de répartition des CV basés sur les densités (CVDT et CVDM) sont tracés (fig. 9 et 10) afin d'apprécier de manière globale le degré de stabilité des peuplements.
Figure 9 : Répartition des CVDT Figure 10 : Répartition des CVDM
Le degré de stabilité peut être évalué en se basant sur la classification proposée par Freeman et al. (1988) :
1) CV ≤ 25% fortement stable
2) 25% < CV ≤ 50% modérément stable 3) 50% < CV ≤ 75% modérément fluctuant 4) CV > 76% fortement fluctuant
La classification proposée par Freeman est applicable aux CV basés sur les densités (CVDM, CVDT et CVDE) (BELKESSAM, 1998). La répartition des CV selon les différentes classes est présentée dans le tableau 2.
Tableau 2 : Effectif et pourcentage de stations selon les valeurs de CV
CV ≤ 25% 25% < CV ≤ 50% 50% < CV ≤ 75% CV > 76% Effectif (sur 473) 1 13 88 370 CVDM Pourcentage 0.2 % 2.8 % 19 % 78 % Effectif (sur 473) 28 192 171 82 CVDT Pourcentage 6 % 40.5 % 36 % 17.5 % CemOA : archive ouverte d'Irstea / Cemagref
Selon cette classification, les valeurs de CVDT correspondent essentiellement à des peuplements modérément stables à modérément fluctuants. La densité totale des peuplements varie donc plutôt de façon modérée. En revanche, les valeurs de CVDM correspondent plutôt à des peuplements fortement fluctuants. Ainsi, les densités des espèces ne sont pas stables dans le temps. Cela suggère qu'à l'échelle globale du peuplement, il y a moins de variation qu'au niveau de la densité spécifique à l'intérieur des peuplements. Au niveau de la densité totale du peuplement, la variation interspécifique se trouve en partie masquée. En moyenne, les espèces varient de façon forte au niveau de leur densité mais l'impact sur la densité globale du peuplement semble limité.
Les quelques études conduites sur les peuplements de poissons de cours d'eau aboutissent à des résultats similaires (BELKESSAM, 1998). Ainsi, Belkessam et Oberdorff (1998) obtiennent, dans une étude sur les peuplements de poissons des cours d'eau bas-normands, un CVDT moyen de 48,49 et un CVDM moyen de 75,33. Hugueny et al. (1995) (BELKESSAM, 1998), dans une étude de la variabilité temporelle de 18 peuplements de poissons de rivières françaises et ouest-africaines, obtiennent des valeurs de coefficients de variations assez proches de ces tendances (CVDT = 71, CVDM = 107,2). De même, Grossman et al. (1990), lors d'une étude portant sur 10 rivières du Midwest des USA, obtiennent des valeurs de CVDM comprises entre 70 et 135 (moyenne = 96). D'autre part, si la variabilité temporelle des poissons est comparée à celle des autres grands groupes d'animaux, il s'avère que, quel que soit le descripteur utilisé, ces derniers sont beaucoup plus stables : CV compris entre 10 et 70 pour les populations d'oiseaux des zones tempérées, et environ 50 pour celles des insectes et des petits mammifères (BELKESSAM, 1998, d'après PIMM, 1991).
Le graphique de répartition du CV basé sur la richesse spécifique locale (CVRSL; fig.11) nous permet d'évaluer la variation de la richesse spécifique des peuplements au cours du temps.
Figure 11 : Répartition des CVRSL
CemOA
: archive
ouverte
d'Irstea
Nous voyons ici que les valeurs de coefficient de variation sont assez faibles. Cela suggère que la richesse spécifique varie peu au sein des peuplements. Ainsi, bien que le nombre d'individus par espèce varie fortement, le nombre d'espèces change peu dans le temps, ce qui ne signifie pas forcément qu'il s'agit des mêmes espèces.
Dans l'étude de Belkessam et Oberdorff (1998), la moyenne des CVRSL est de 13,68, avec des valeurs qui sont toutes du même ordre de grandeur que celles obtenues dans notre étude.
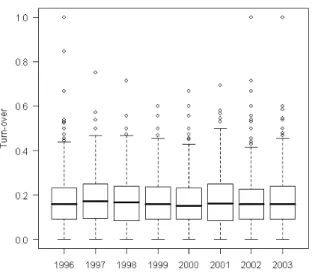
Une vision plus précise des variations temporelles des espèces présentes est apportée par l'analyse des turn-over. Les valeurs observées (fig.12) montrent que dans la grande majorité des cas, le nombre d'espèces changeant d'une année sur l'autre reste inférieur à 20%. D'autre part, le graphique (fig.13) montre, confirmé par un test ANOVA (p-value = 0,29), qu'en moyenne les turn-over restent stables dans le temps même si les conditions hydro-climatiques ont varié plus ou moins fortement d'une année sur l'autre au cours du suivi.
Figure 12 : Répartition des turn-over Figure 13 : Turn-over en fonction de l'année
Ainsi, aucune réponse particulière n'est détectée au niveau des turn-over à des événements climatiques remarquables comme la tempête de 1999 ou la canicule de 2003 (même si pour ce dernier cas certains échantillons de 2003 étaient antérieurs à la période de canicule).
CemOA
: archive
ouverte
d'Irstea
- Influence de la technique d'échantillonnage sur la variabilité des peuplements
Comme expliqué plus haut, deux méthodes d'échantillonnage ont été employées selon les caractéristiques du cours d'eau considéré. Dans un cas l'échantillonnage a été réalisé à pied, dans un autre cas en bateau. Il est probable que cette différence ait des conséquences sur les observations. A titre d'exemple, la variation du CVDM selon la méthode d'échantillonnage est représentée sur le graphique suivant (fig.14).
Figure 14 : CVDM selon la méthode d'échantillonnage
Lorsque l'échantillonnage est effectué par bateau les CV prennent des valeurs plus importantes. Cette tendance est confirmée par des tests ANOVA qui montrent que CVDM et CVDT sont plus élevés avec l'échantillonnage par bateau qu'avec celui à pied. Le graphique du CVRSL montre une tendance identique mais moins accentuée et le test ANOVA donne une probabilité critique non significative à un seuil de 5 %. Nous pouvons conclure de ces résultats que la technique d'échantillonnage employée n'est sans doute pas négligeable et peut induire un biais dans nos résultats. Nous avons la confirmation que la pêche en bateau partielle le long des rives donne une image imparfaite du peuplement en place (OBERDORFF et al., 2002, d'après CHANCEREL, 1992) mais une bonne représentation de la richesse en espèces puisque la technique d'échantillonnage n'a pas d'effet sur le CV basé sur la richesse spécifique, CVRSL. La technique d'échantillonnage ne sera pas utilisée dans les tests suivants car beaucoup de données sont manquantes et cela conduirait à exclure un grand nombre de stations des tests. Ce paramètre sera remplacé par le ratio échantillonné pour lequel nous disposons de toutes les données. Le ratio échantillonné englobe également l'aspect de la technique d'échantillonnage puisque les stations qui n'ont pas été échantillonnées complètement correspondent le plus souvent à celles échantillonnées par bateau.
CemOA
: archive
ouverte
d'Irstea
- Variabilité des peuplements en fonction des caractéristiques des stations et de l'échantillonnage
Pour chaque paramètre étudié, un graphique est tracé afin d'observer les variations du CV ou du T moyen selon ce paramètre. Ensuite, un test de régression ou une ANOVA est réalisé pour savoir s'il existe une relation entre le CV ou le T moyen et ce paramètre. Les tableaux suivants (tableaux 3 et 4) recensent les paramètres étudiés et les résultats obtenus. Le seuil des tests est fixé à 5%.
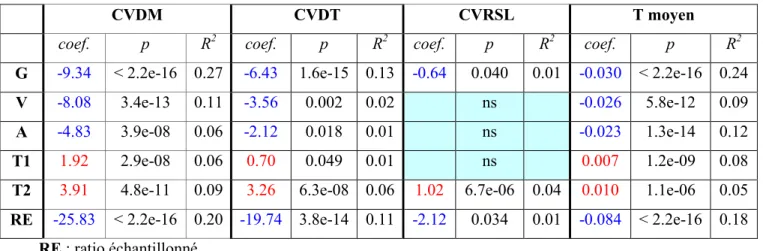
Tableau 3 : Résultats des régressions simples pour les variables quantitatives
CVDM CVDT CVRSL T moyen
coef. p R2 coef. p R2 coef. p R2 coef. p R2
G -9.34 < 2.2e-16 0.27 -6.43 1.6e-15 0.13 -0.64 0.040 0.01 -0.030 < 2.2e-16 0.24
V -8.08 3.4e-13 0.11 -3.56 0.002 0.02 ns -0.026 5.8e-12 0.09
A -4.83 3.9e-08 0.06 -2.12 0.018 0.01 ns -0.023 1.3e-14 0.12
T1 1.92 2.9e-08 0.06 0.70 0.049 0.01 ns 0.007 1.2e-09 0.08
T2 3.91 4.8e-11 0.09 3.26 6.3e-08 0.06 1.02 6.7e-06 0.04 0.010 1.1e-06 0.05
RE -25.83 < 2.2e-16 0.20 -19.74 3.8e-14 0.11 -2.12 0.034 0.01 -0.084 < 2.2e-16 0.18
RE : ratio échantillonné
Tableau 4 : Résultats des ANOVA pour les variables qualitatives
CVDM CVDT CVRSL T moyen
p-value p-value p-value p-value
Etat de l'habitat < 2.2e-16 2.427e-06 0.002274 < 2.2e-16
Qualité de l'eau 0.01338 ns ns ns
HER groupes 1.157e-13 6.366e-05 0.001466 < 2.2e-16
Les tableaux montrent que les résultats pour CVDM, CVDT et T moyen sont globalement similaires alors que CVRSL ne montre pas souvent de relation significative avec les paramètres.
Les résultats des ANOVA concernant l'état de l'habitat et la qualité de l'eau sont illustrés par les graphiques suivants (fig.15 et 16).
CemOA
: archive
ouverte
d'Irstea
Figure 15 et 16 : CVDM selon la qualité de l'habitat et selon la qualité de l'eau
Ces premiers résultats indiquent que tous les paramètres semblent liés à la variabilité à l'exception de la qualité de l'eau. Ainsi, l'échantillonnage, l'environnement naturel et les perturbations anthropiques (via l'état d'altération de l'habitat) semblent tous susceptibles d'expliquer une part de la variabilité observée. Il est difficile à ce stade de savoir si les résultats obtenus résultent de relations réelles ou s'il s'agit au moins pour partie d'artéfacts dans la mesure où la plupart des variables explicatives sont plus ou moins fortement corrélées entre elles (à titre d'exemple, il existe une relation forte entre le ratio échantillonné et la taille du milieu, les petits cours d'eau étant échantillonnés complètements alors que, sur les grands cours d'eau, l'échantillonnage ne concerne qu'une partie des rives). Dans ces conditions, il apparaît clairement à la suite de ces résultats que des analyses facteur par facteur ne sont pas pleinement satisfaisantes.
Une seconde série d'analyses est donc effectuée à l'aide de régressions multiples qui ne retiennent que les facteurs les plus explicatifs et limitent les artéfacts liés à l'existence d'auto-corrélations entre variables explicatives.
Il est important de noter que ces analyses n'ont pas pour objectif de fournir des modèles prédictifs mais simplement de voir ce qui peut expliquer les variations observées. Les résultats de l'analyse sont synthétisés dans le tableau suivant (tableau 5).
CemOA
: archive
ouverte
d'Irstea
Tableau 5 : Synthèse des résultats des régressions multiples
CVDM CVDT CVRSL T moyen
coef. p-value coef. p-value coef. p-value coef. p-value
G -2.979 0.012 -6.796 2.3e-09 -0.008 0.034
V -3.965 0.000
A -0.987 0.027 -0.009 0.017
T1 1.175 0.000 -0.306 0.041 0.003 0.023
T2 2.278 8.3e-05 2.412 5.3e-06 1.027 8.0e-05 0.006 0.017
Ratio échantillonné -12.715 0.000 -7.181 0.042 -0.025 0.048 Nul Faible -0.086 0.980 0.412 0.795 0.011 0.382 Interm 8.900 4.8e-4 3.334 0.005 0.029 0.001 Altération de l'habitat
Fort 8.217 9.8e-4 1.644 0.142 0.038 2.1e-5
Faible Moyen -2.813 0.004 Altération de l'eau Fort 1.777 0.324 Gpe1 Gpe2 -1.782 0.603 -0.007 0.559 Gpe3 19.196 0.006 -0.003 0.886 Gpe4 1.058 0.787 -0.028 0.038 Gpe5 0.546 0.933 -0.052 0.027 HER groupes Gpe6 1.544 0.793 -0.019 0.327 R2 0,396 0,307 0,087 0,383
Au vue de ces résultats, nous pouvons conclure que la variabilité des peuplements échantillonnés par pêche électrique dépend à la fois de l'échantillonnage, de l'environnement naturel et des altérations anthropiques (essentiellement de la qualité d'habitat).
Les résultats indiquent que la stabilité des peuplements piscicoles (vue au travers les échantillonnages par pêche électrique) diminue de l'amont vers l'aval. En effet, lorsque la station est située plus en amont (G augmente), la variabilité des peuplements diminue. Ce résultat va à l'encontre des théories développées dans la littérature (SCHLOSSER, 1990) qui attribuent une plus grande variabilité temporelle des peuplements en amont, où les conditions environnementales sont soumises à de plus importants changements.
Le paramètre de vitesse d'écoulement n'est significatif que pour expliquer les variations du
CemOA
: archive
ouverte
d'Irstea
d'écoulement est plus importante. Ce paramètre est difficile à analyser car il correspond à des situations naturelles. Or, en pratique, beaucoup de rivières sont altérées et comportent fréquemment des seuils ou des barrages qui modifient le régime hydrologique.
Le résultat pour l'altitude suggère que les peuplements sont plus stables en haute altitude. Or il se trouve que peu d'espèces vivent dans les stations d'altitude élevée ce qui pourrait expliquer ce résultat, d'autant plus qu'il porte sur CVRSL et T moyen qui sont les paramètres qui se basent essentiellement sur la variabilité des espèces.
L'indicateur de la température de l'air T1 ne donne pas des résultats similaires avec tous les paramètres de variation. Il semble se dégager en priorité la conclusion que lorsque la température de l'air augmente les peuplements varient davantage. C'est cependant l'inverse qui est observé avec CVRSL.
En revanche le paramètre d'amplitude thermique T2 est considéré comme facteur explicatif dans le modèle dans tous les cas. Les peuplements varient plus lorsque l'amplitude thermique est plus importante. Il s'agit ici d'une amplitude thermique saisonnière qui indique par exemple si l'année est plus ou moins favorable à la reproduction. Nous pouvons émettre l'hypothèse que les milieux avec une forte valeur de T2 (fort contraste saisonnier) sont aussi ceux qui devraient présenter les plus fortes variations interannuelles de température d'où des plus fortes variations de peuplements. En effet, nous pouvons supposer que lorsque l'amplitude thermique est plus importante, les conditions environnementales varient entraînant une fuite des poissons si elles deviennent mauvaises ou au contraire un arrivage si elles deviennent meilleures. Cela aura également probablement un impact sur les succès de reproduction. Des données sur la variation thermique pendant toute la durée du suivi auraient permis des analyses plus pertinentes.
Nous voyons aussi que lorsque le ratio échantillonné augmente, ce qui correspond aux petits milieux, la variabilité des peuplements diminue. Bien que l'aspect échantillonnage ne soit pas indépendant de la taille du milieu donc du gradient longitudinal, nous voyons grâce à la régression multiple que chacun explique une part différente des variations. Il existe bien un biais introduit par l'échantillonnage, qui diminue lorsque l'échantillonnage est plus complet. Aucun effet de l'échantillonnage sur le CVRSL n'est montré. Cela s'explique de la même manière que pour l'analyse selon la méthode d'échantillonnage réalisée précédemment, par
CemOA
: archive
ouverte
d'Irstea
une bonne représentation de la richesse en espèces même dans les grands cours d'eau où la pêche s'effectue en bateau, contrairement à la structure du peuplement qui est moins bien évaluée dans ce cas.
Le logiciel R permet de détailler l'effet de chaque groupe d'hydro-écorégions (HER). Les résultats montrent, pour le CVDM, que les peuplements des stations situées dans les régions du groupe 3 (formations périphériques des massifs) varient plus que les autres. Pour le T moyen, ce sont les peuplements des stations des groupes 4 et 5 (massifs anciens et massifs récents) qui varient moins que les autres. Les graphiques correspondant aux tests ANOVA avec CVDM et avec T moyen sont les suivants (fig.17 et 18) :
Figure 17 : CVDM selon les groupes d'HER Figure 18 : T moyen selon les groupes d'HER
Ces résultats sont difficiles à expliquer pour l'instant. Ils ne correspondent pas à ce qui aurait pu être attendu, à savoir une plus grande stabilité dans les plaines.
Concernant l'état de l'habitat, l'analyse montre une augmentation de la variabilité lorsque la dégradation augmente. Ainsi, les états de dégradation sont classés dans l'analyse comme ayant un effet croissant sur la variabilité (nul<faible<intermédiaire<fort), comme nous le voyons sur la figure 15. Ce paramètre a été évalué essentiellement par rapport au niveau de recalibrage des cours d'eau. Lorsque le cours d'eau a été fortement recalibré, le milieu est beaucoup moins diversifié et, lorsqu'une variation de débit se produit, les conditions se modifient fortement dans la station sans possibilité de zones refuges pour les poissons. Au contraire, dans les milieux peu ou pas impactés par l'homme, une mosaïque diversifiée d'habitats se maintien et
CemOA
: archive
ouverte
d'Irstea
donc entraîner une augmentation de la variabilité des peuplements car les poissons vont devoir se déplacer pour chercher ailleurs des refuges. Il est dommage de ne pas disposer pour l'instant de données sur l'hydrologie qui auraient permis de prendre en compte les variations de débit, les crues et les étiages.
La qualité de l'eau n'est pas considérée comme un facteur explicatif sauf pour CVRSL. Le résultat obtenu indique que les peuplements varient plus lorsque la qualité d'eau est moyenne. Ce résultat est difficile à expliquer, mais il faut tenir compte du fait que la part de variance expliquée par le modèle pour CVRSL est beaucoup plus faible que pour CVDM, CVDT et T moyen.
Analyse de la variabilité des populations et des différences entre espèces
- Variations du CVDE selon les caractéristiques des espèces
Cette partie a pour but de mettre en évidence les relations qui existent entre la variabilité observée à l'échelle des populations et les caractéristiques biologiques et écologiques des espèces. Compte-tenu des modalités de calcul des CVDE, dix-huit espèces sont exclues des analyses car elles ne sont jamais présentes neuf années consécutives (toutes ces espèces sont des espèces plutôt rares sur le territoire français). Les liens entre CVDE et les caractéristiques des espèces sont testés avec des régressions simples ou des ANOVA dont le seuil est fixé à 5%. Néanmoins, compte tenu du nombre d'observations relativement faible (39 espèces), les cas où la probabilité critique est inférieure à 10 % sont également présentés. Les résultats obtenus figurent dans le tableau suivant (tableau 6). Les paramètres pour lesquels aucune tendance particulière n'a été montrée ne sont pas représentés.
Tableau 6 : Résultats synthétiques des tests sur les caractéristiques des espèces
Paramètre Coef. p-value R2
Taille maximale connue (log) -20.453 0.003 0.24 Relation
significative au
seuil 5% Fécondité (log) -4.280 0.040 0.13
Age à maturité -2.763 0.063 0.60
Longévité -0.698 0.054 0.11
Fécondité relative (log) 7.871 0.076 0.12 Relation
significative au seuil 10%
Guilde trophique de l'adulte x 0.099
CemOA
: archive
ouverte
d'Irstea
Les graphiques suivants (fig.19 et 20) illustrent certains résultats. La droite de régression apparaît en rouge.
Figure 19 et 20 : CVDE selon la taille maximale connue (log) et selon la fécondité (log)
Le paramètre "guilde trophique de l'adulte" est un facteur qualitatif. Le nombre d'espèces dans certaines classes étant très faible, il est difficile de tirer des conclusions sur ce paramètre. Le graphique suivant (fig.21) illustre la variation des CVDE selon la guilde trophique de l'adulte.
Légende
HERB : herbivore INSV : insectivore OMNI : omnivore PISC : piscivore
Figure 21 : CVDE selon la guilde trophique de l'adulte
Seule la différence entre les classes HERB et PISC donne une valeur inférieure au seuil 10%. Cependant, seules deux espèces (la bouvière et le hotu) appartiennent à la guilde des herbivores, ce qui est insuffisant pour avoir un résultat fiable. Par conséquent, aucune conclusion très claire ne peut être tirée concernant ce facteur.
CemOA
: archive
ouverte
d'Irstea
Les autres résultats indiquent que les espèces de grande taille ont une plus faible variabilité que celles de petite taille. Les espèces qui vivent plus longtemps, qui ont généralement un âge à la maturité plus élevé, semblent aussi plus stables que les autres. Une forte fécondité correspond à des espèces plus stables, par contre une forte fécondité relative semble correspondre à des espèces moins stables. Ces résultats peuvent sembler contradictoires mais ils reflètent simplement la chose suivante : les "gros" poissons pondent plus d'œufs que les "petits" mais, ramené au poids, ce sont les "petits" poissons qui pondent plus que les "gros". Il semble donc que, globalement, les "gros" poissons présentent des populations plus stables que les "petites" espèces. Lorsque les résultats obtenus pour la taille, l'âge et la fécondité sont regroupés, il apparaît qu'ils sont en concordance avec un type de stratégie biologique : les espèces de poissons de grande taille, qui vivent longtemps et pondent beaucoup d'œufs sont plus stables que les autres.
Une régression multiple a également été réalisée pour tenter d'évaluer les effets conjoints des différentes variables explicatives. Seule la taille maximale connue ressort dans le modèle final ce qui pourrait s'expliquer par les fortes dépendances qui existent entre les différents paramètres décrivant la biologie et l'écologie des espèces. Ce résultat est en accord avec les conclusions tirées ci-dessus, car la taille maximale connue englobe plusieurs caractéristiques correspondant à une stratégie biologique.
- Variabilité d'une espèce selon sa position vis-à-vis de son optimum écologique
Le but de cette partie est de voir s'il existe un lien entre la position d'une espèce dans sa niche écologique et la variabilité de ses populations. Seuls les cas où l'espèce est présente les neuf années consécutives sur la station sont sélectionnés. Les probabilités de présence des espèces par station ont été calculées à partir des modèles développés dans le cadre de l'IPR. Ces probabilités sont utilisées comme un indicateur de la situation de l'espèce par rapport au centre de sa niche écologique. Comme les probabilités de présence ne peuvent pas être calculées pour toutes les espèces (34 espèces prises en compte) et que certaines espèces ne sont jamais présentes neuf années dans une station, les tests ne portent que sur 15 espèces. Les tests effectués sont des régressions linéaires simples, basées sur les CVSP. Compte tenu du faible nombre de valeurs, le seuil est fixé à 10%. Les résultats sont présentés dans le tableau 7. Les espèces pour lesquelles aucune tendance n'a été montrée ne figurent pas dans ce tableau. CemOA : archive ouverte d'Irstea / Cemagref
Tableau 7 : Résultat des tests en fonction des probabilités de présence
Espèce Pente p-value R2
Chabot -22.213 0.051 0.02 Gardon 29.781 0.001 0.05 Goujon 32.952 0.007 0.03 Loche franche -19.77 0.057 0.02 Perche -26.000 0.012 0.06 Truite -42.79 0.002 0.05 Vairon -34.331 0.006 0.04
Nous nous attendons à observer une plus grande variabilité lorsque la probabilité de présence (pp) de l'espèce est faible. Cela signifierait que les populations sont plus stables dans des conditions d'environnement optimales. C'est effectivement ce qui est observé pour 5 espèces ici : le chabot, la loche franche, la perche, la truite et le vairon. Nous voyons que plus les probabilités de présence de ces espèces augmentent, plus les CVSP sont faibles et donc plus les populations sont stables.
Figure 22 : CVSP selon la pp de la perche
A titre d'exemple, le graphique (fig.22) illustre le cas de la perche.
Le résultat inverse est observé pour le gardon et le goujon (fig.23). Cela peut être expliqué par des facteurs qui sont propres à ces espèces. Le gardon par exemple fait l'objet de beaucoup de repeuplements pas l'homme (environ 2000 tonnes par an; KEITH, 2001), ce qui influe très probablement sur la variabilité de ses populations. Le goujon est connu pour avoir une densité de population qui fluctue considérablement d'une année sur l'autre (KEITH, 2001), ce qui pourrait expliquer les fortes valeurs de CV observées dans certains cas chez cette espèce.
CemOA
: archive
ouverte
d'Irstea
CONCLUSION
L'un des objectifs de ce début de stage était de quantifier la variabilité des peuplements de poissons des cours d'eau français et de la mettre en relation avec différents facteurs potentiellement explicatifs : l'environnement naturel, les perturbations anthropiques et les aspects liés à l'échantillonnage. Les analyses ont montré que la variabilité des peuplements, appréhendée à travers un échantillonnage par pêche électrique, dépend de ces trois types de facteurs. Les facteurs environnementaux impliqués sont principalement le gradient longitudinal des cours d'eau et l'amplitude thermique saisonnière. Les perturbations d'origine anthropique augmentent la variabilité des peuplements, à travers l'altération de l'habitat des poissons (recalibrage des cours d'eau). Enfin nous avons vu que la méthode d'échantillonnage introduit également une part de variabilité dans les résultats observés.
Un second objectif consistait à étudier la variabilité des populations de poissons en fonction des caractéristiques biologiques et écologiques des espèces. Les analyses ont montré que les espèces de grande taille, à forte longévité et forte fécondité ont des populations plus stables. Enfin, nous avons testé si les populations d'une espèce donnée sont plus stables lorsque l'espèce est proche du centre de sa niche écologique. Nous avons pu montrer que c'était effectivement le cas pour certaines espèces.
Ces résultats apportent des informations intéressantes sur les peuplements de poissons des cours d'eau français, parfois contraires à ce qui était attendu, comme par exemple l'effet inverse du gradient longitudinal sur la variabilité des peuplements par rapport aux données de la littérature. L'analyse des résultats va devoir être approfondie de manière à rassembler des éléments explicatifs.
Dans la suite de ce stage, ces questions seront probablement étudiées de nouveau à l'aide de paramètres supplémentaires (régime hydrologique, nouvelles données sur la qualité de l'eau…). En outre, à une échelle plus fine, les analyses vont porter plus précisément sur l'évaluation des phénomènes d'extinction et de colonisation locales, en tentant de les relier à différents facteurs explicatifs. Les méthodologies s'appuieront sur des démarches déjà développées dans le cadre de travaux ayant abordés ce type de problématique appliquée aux peuplements de poissons (GOTTELI et TAYLOR, 1999, OBERDORFF, HUGUENY et VIGNERON, 2001). CemOA : archive ouverte d'Irstea / Cemagref