[
[
[
[
[
[
l
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE DE L'ENSEIGNEMENT SURERIURE ET
DE LA RECHERCHE SCIENTIFIQUE
Université de flJEL
Faculté des Sciences
Mémoire
de fin d'études présenté en vue de l'obtention du diplôme des
études
Supérieur
(D.E.S) en·Biologie
Option : Biologie et physiologie végétale
Thème
Effet du déficit hydrique sur l'accumulation de
certains composés organiques
·
:./'~~
1
'
4'0\ '\
1J;
\.
.
.
Session : juin 2008Réalisé par :
Abdelmoula Rabiaà
Medoukali Rebiha
:Remerciement
Nous remercions tout d'abord Dieu qui nous
·
a donnée le courage et la santé durant
les années d'études.
Nous remercionst en première lieu notre encadreur :
MlleBenterouche
1.pour son
aide et pour la confiance qu'elle a bien voulus mettre en nous, et également pour ces
conseils tout au long de ce modeste travail.
Nous tenons a remercie également les nombres du jury, pour avoir accepter de juger
ce mémoire
.
Nous ne terminons pas sans avoir exprimer nos vifs remerciements à tout les
enseignants qui ont contribué à notre formation.
Enfin nous remercions touts les personnes qui nous ont aidé de prés ou de loin même
par le simple mot d'encouragement.
Sommaire
Introduction ... 01
Chapitre 1: L'eau et la plante
I.1. Besoins des plantes en eau ... · ... 03I.2. Rôle de l'eau dans la plante ... 05
I.3. Caractéristique de l'état hydrique du sol et de la plante ... 06
I.3.1. Dans le sol ... 06
I.3.1.1. La capacité au champ (CC) ... 06
I.3.1.2. Le point de flétrissement permanent (PFP) ... 06
I.3 .1.3 L'eau utilisable ou réserve utile (RU) ... 06
I.3.1.4. La teneur en eau optimal ... 07
I.3.2. Dans la plante ... 07
I.3.2.1. La teneur en eau ... 07
I.3.2.2.La teneur relative en eau ou contenu relatif en eau (RWC) ... 08
I.3 .2.3 .Le potentiel hydrique 'l'h et ses composantes ... 08
I.4. L'absorption de l'eau par les plantes ... 09
Chapitre II : Le déficit hydrique
II. l. Notion de sécheresse et de déficit hydrique ... 13II.2. Mécanismes d'adaptation des plantes au déficit hydrique ... 13
II.2.1. Esquive ou échappement ... 14
II.2.2 Evitement ... 14
II.2.3. La tolérance ... 14
II.2.4.Acclimatation ... 16
II.3.3. Effets du déficit hydrique sur le rendement et le développement ... 19
II.3.4. Effet du déficit hydrique sur les paramètres du statut hydrique ... 19
II.3.4.1.Sur la teneur relative en eau ... 19
II.3.4.2. Sur le potentiel hydrique foliaire ... 21
II.3.5. Effets du déficit hydrique sur les paramètres physiologiques ... 22
II.3.5.1. Effets sur la réponse stomatique ... 22
11.3.5.2. Effets sur la structure membranaire ... 24
II.3.5.3. Effets sur la photosynthèse ... 24
Chapitre III: Effet du déficit hydrique sur l'accumulation des Composées organiques IIl.1. Effets du déficit hydrique sur le métabolisme protéique ... 25
111.1.1. La proline ... 2
IIl.1.1.1. Voies de synthèse de la proline ... 28
IIl.1.1.2.Synthese de la proline par autre voie ... 30
III.1.2. Les protéines LEAS (late embryogenèses arbundant) ... 31
III.2. Effets du déficit hydrique sur le métabolisme glucidique ... 32
III-3- Effets du déficit hydrique sur le métabolisme hormonale ... 33
Liste âes tafJ{eaux
Tableau n°1 : Valeurs moyennes des teneurs en eau des différents organes des plantes.
Tableau n°2: Besoins totaux en eau de quelques plantes (pluies plus irrigation) en litres par kilo de matière sèche.
Tableau n°3: Paramètre d'adaptation à la sécherreses.
Tableau n° 4: Influence d'un choc osmotique sur les acides aminés libres et autres composés positifs à la ninhydrine des feuilles de cotonnier.
Liste âes ftnures
Figure n°1: Caractéristique de l'état hydrique du sol.
Figure n°2: Poil absorbant.
Figure n°3: Mycorhizes.
Figure n°4: Coupe transversale d'une racine.
Figure n°5: La plante et son environnement hydrique
Figure n°6 : Les principaux effets du déficit hydrique sur les composantes du rendement Figure n° 7: La perte de turgescence chez les cellules
Figure n°8: Les stomates.
Figure n°9: Diffusion de la vapeur d'eau par les stomates ouverts
Figure n°10: Fermeture stomatique et diminution de la transpiration foliaires (Stratégie d'évitement)
Figure n°11 : Voie de synthèse de la proline à partir du glutamate. Figure n°12 : Synthèse de la proline par d'autre voie
'l'n : Potentiel hydrique. ABA: Acide abscissique CC : Capacité au champ
Liste tfes abréviations
LEAS : Late Embryogenesis Arbundantto MF: Matière fraîche
MS: Matière sèche
PEP: Point de flétrissement permanent. RU: Réserve utile.
RWC: (Relative Water Content) la teneur relative en eau. Turg: Turgescent
TE: Teneur en eau
I I I 1 Introduction
Introauction
L'eau est indispensable à toute vie. Elle est le constituant le plus important des végétaux; elle forme habituellement environ
60%
de leur poids frais. Milieu dans lequel a lieu la quasi-totalité des processus biologiques, elle joue un rôle de transporteur, de solvant, d'agent de réaction chimique; elle donne aux plantes leur turgescence; grâce à la chaleur latente élevée de vaporisation de l'eau, la transpiration permet aux plantes de perdre une partie importante de l'énergie qu'elles reçoivent du soleil et de supporter ainsi son rayonnement de façon continue sans pour autant subir un échauffement excessif. L'eau est donc impliquée tant dans les structures que dans les processus de transport et le métabolisme des végétaux.De ce fait, la disponibilité de l'eau pour la croissance des plantes est un facteur limitant important. Dans les régions sèches, elle constitue la principale entrave à la croissance et au développement des plantes. Dans les zones tempérées, des périodes de sécheresse inattendues peuvent également devenir, en l'absence de possibilités d'irrigation, un facteur crucial de réduction des productions végétales.
Contrairement aux animaux qui peuvent se déplacer lorsque les conditions de
\
deviennent défavorables, Ues plantes ont développé des stratégies d'adaptation fondées le contrôle et l'ajustement de leurs systèmes
métabolique~
vie sur
En effet, les plantes ont adopté différentes stratégies pour survivre en situation de stress hydrique dont les principales consistent à :
- Echapper au stress hydrique: la plante effectue son cycle de reproduction en dehors des périodes critiques,
\ , . - /
- Eviter le stress hydrique: la plante active des mécanismes qui permettent de maintenir un potentiel hydrique élevé, ceci en augmentant les capacités d'absorption de l'eau et/ou en réduisant les pertes. Ainsi pour mieux exploiter les réserves hydriques du sol, la plante va augmenter la profondeur de son système racinaire, sa densité et son degré de ramification. De plus, la fermeture des stomates, la diminution des surfaces évaporantes par enroulement foliaire, la sénescence et l'abscission précoces des feuilles sont autant de moyens pour limiter les pertes en eau.
/ '
- f
- Tolérer le stress hydrique: la plante fait appel à un processus de maintien osmotique : le maintien d'une turgescence positive est rendue possible par l'accumulation de solutés particuliers, plusieurs types de molécules peuvent assurer l'ajustement osmotique. Les sucres solubles et les acides aminés, en particulier la proline, sont les composants majeurs de cet '-
~
, ) ~ I I I I I ~ Il
Chaoitre l
:L
'eau
et la Dlante
I .1 Besoins iles plantes -en -eail :
L'eau couvre 70% de la surface du globe et constitue 85 à 90% de la matière vivante animale et végétale (Laval-Martin et Mazliak, 1995). Elle constitue par l'intermédiaire des solutions du sol la base de l'alimentation de la plante.
Tous les organes de la plante sont très riches en eau, elle est très souvent entre 70 et 90% de leur masse (Tourte et al, 2005). Mais chez les algues elle atteint jusqu'à 90% de leur masse total (Zella et Smadhi, 2006)
L'eau représente plus de 70% de la masse des parties non lignifiées de la plante. Bien que certaines plantes tolérantes à la dessiccation puissent ne contenir que 20% d'eau et que les graines sèches n'en contiennent que 5%, ces deux types d'organismes sont métaboliquement inactifs et la reprise d'une activité métabolique importante est conditionnée par la restauration d'un contenu normal en eau (Hopkins, 2003).
Il existe différentes formes d'eau dans la plante. On trouvera l'eau de constitution qui donne la conformation des molécules, l'eau d'imbibition qui maintient les molécules en solution et l'eau capillaire piégée dans les interstices et espaces intercellulaires.
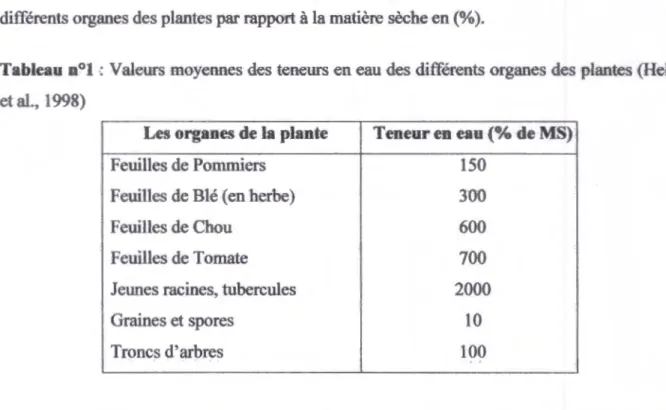
Chez les plantes on trouve un contenu en eau variable selon les plantes et l'organe considéré. Le tableau suivant présente quelques valeurs moyennes des teneurs en eau des différents organes des plantes par rapport à la matière sèche en (% ).
Tableau n~l : Valeurs moyennes des teneurs en eau des différents organes des plantes (Heller et al., 1998)
Les organes de la plante Teneur en eau (% de MS)
Feuilles de Pommiers 150
Feuilles de Blé (en herbe)
JOO
Feuilles de Chou 600
Feuilles de Tomate 700
Jeunes racines, tubercules 2000
Graines et spores 10
Troncs d'arbres 100 ..
Le contenu réel en eau dépend également du tissu et du type cellulaire, ·et ·des conditions physiologiques et environnementales.
t_
teneur en eau diminue aussi avec l'age,Chapitre I : L 'eau et la plante c'est-à-dire les tissus jeunes en voies de croissance sont plus riches en eaux que les tissues âgées.
Dans les cellules adultes l'eau est située pour la plus grande part dans la vacuole qui occupe souvent plus de 95% du volume cellulaire. Mais le cytoplasme est lui-même très hydraté. Les cellules jeunes sont aussi très riches en eau, en effet, la teneur en eau peut atteindre jusqu'à 2000 % de sa matière sèche (Heller et al., 1998).
Les mesures montrent qu'il faut en général environ 300L d'eau pour l'élaboration d'lg de matière sèche. Selon Zella et Smadhi (2006), le maïs à besoin de 250L d'eau pour fabriquer 1 Kg de matière sèche.
Le tableau suivant, montre les besoins de quelques cultures en eau pour l'élaboration d'un kilogramme de matière sèche
Tableau n<>i: Besoins totaux en eau de quelques plantes (pluies plus irrigation) en litres par kilo de matière sèche (Anonyme, 2006)
Cultures Litres d'eau
Maïs fourrage* 238 Banane 346 Maïs grain* 454 Orge* 524 Pomme de terre* 590 Blé* 590 Soja 900 Riz pluvial 1600 Riz inondé 5000
(NB: *en zones tempérées)
D'une manière générale selon la FAO le besoin en eau d'une culture est la quantité d'eau nécessaire à couvrir les pertes d'eau par évapotranspiration (Agrab, 2005)
Chapitre I : L 'eau et la olante
1.2. Rôle de l'eau dans la plante :
L'eau est nécessaire à la plante comme tout être vivant et à plusieurs niveaux:
1.2.1. La turgescence cellulaire:
L'eau contribue au maintien de la cellule et par la même des tissus et des organes (Laberch, 1999), en effet, elle donne une grande rigidité aux tissus végétaux et assure le port dressé des organes dépourvus de tissu de soutien. Quand la plante subit une alimentation hydrique insuffisante, les cellules perdent de l'eau et le volume de la vacuole diminue (plasmolyse). Aussi la première manifestation visuelle d'une déshydratation se traduit par la perte de turgescence qui conduit à un flétrissement et à l'abandon du port érigé (Adjal, 2006).
1.2.2. Le mouvement d'organes et de cellules
Elle commonde divers mouvement d'organes (feuilles, étamines) et de cellules (stomates) (Heller et al., 1998) et permet le déplacement des anthérozoïdes chez les thallophytes, les ptéridophytes,les bryophytes et les pré spermaphytes (Laberche, 1999).
1.2.3. Source d'éléments essentiels
L'eau possède également d'excellentes propriétés solvantes, ce qui permet l'absorption et
....
la distribution des nutriments minéraux et autres solutés nécessaires à la croissance (Hopkins, 2003) et parallèlement il véhicule ces nutriments nécessaires pour le métabolisme (Laberche, 1999).
1.2.4. Source d'énergie
La transparence de l'eau aux radiations lumineuses visibles permet aux radiations solaires de pénétrer les milieux aqueux des cellules, où elles fournissent l'énergie nécessaire à la photosynthèse (Hopkins, 2003).
1.2.5. Régulation thermique
L'eau permet de réguler la température des parties aériennes et aide la plante à dissiper la chaleur absorbée sous forme de radiations solaires par le phénomène de transpiration.
Pour une plante, aussi importante que soit la teneur interne en eau, cette dernière ne présente , en fait qu'une très faible partie de la quantité absorbée par les racines (environ 1 % selon Morard, (1995) in Adjel, (2006). En effet, l'eau n'est pas a proprement parler consommée. Elle ne fait que passer du sol à l'atmosphère avec un flux très important comme,
Chapitre I : L 'eau et la plante
par exemple, 6000 m3 /ha/an pour une culture couvrant le sol en été. Cette eau est émise sous forme de vapeur d'eau (Adjel, 2006).
1.3. Caractéristiques de l'état hydrique du sol et de la plante 1.3.1. Dans le sol
1.3.1.1. La capacité au champ (CC) :
C'est la quantité d'eau qui reste dans un sol égoutté normalement après saturation. A la capacité au champ, l'eau est retenue dans le sol à l'encontre des forces de gravité.
1.3.1.2. Le point de flétrissement permanent (PFP):
Est le taux d'humidité du sol au moment où la plante ne peut plus s y approvisionner en eau (car il est retenu par des forces supérieures à la succion des racines) et commence à souffrir de la sécheresse. Selon Simon (1996), le point de flétrissement est atteint après épuisement de l'eau de capillarité.
1.3.1.3 L'eau utilisable ou réserve utile (RU}!
C'est la différence entre la teneur en eau du sol à la capacité au champ et la teneur en eau du sol au point de flétrissement.
RU=CC-PEP
Au point de flétrissement, une certaine quantité d'eau est présente, mais elle n'est pas disponible à la plante. .. -...
~---
...
.
··•·
.
.. ..
·
...
L ~'
Chapitre I: L'eau et la plante
La texture du sol a une influence directe sur les taux d'humidité à la capacité au champ et au point de flétrissement (et par conséquent sur la RU), comme le montre la figure n°1.
Les sols à texture argileuse ont une forte capacité au champ mais leur point de flétrissement est élevé (emmagasinent davantage d'eau mais une grande partie de ces réserves en eau restent indisponibles pour les plantes). Inversement, les sols sableux, qui
n'emmagasinent que peu d'eau, n'en retiennent qu'une faible partie sous forme hygroscopique. Ce sont là encore les textures équilibrées qui offrent le plus d'avantages
(Simon, 1996).
1.3.1.4. La teneur en eau optimal:
C'est la teneur en eau du sol qui, maintenue durant la période de croissance de la plante,
permet d'obtenir un rendement maximal de la culture. En générale, elle se situe, selon le type de culture, près de la capacité au champ.
1.3.2. Dans la plante
En condition de sécheresse, le sol se dessèche progressivement et les forces qui lient l'eau aux particules du sol deviennent de plus en plus importantes, ce qui rend le passage de l'eau de la solution du sol vers le système racinaire de la plante très difficile. Ceci se reflète automatiquement dans le statut hydrique de la plante qui constitue un bon indicateur de la contrainte hydrique au cours du temps.
Les paramètres d'évaluation du statut hydrique de la plante au cours du temps sont nombreux parmi lesquelles on peut citer la teneur en eau (TE), la teneur relative en eau (RWC)
et le potentiel hydrique (\}'h) et ses composantes.
1.3.2.1. La teneur en eau:
La teneur en eau est exprimée par le poids d'eau en rapport avec le poids de la matière sèche ou de matière fraîche
TE= Pr-Ps I Ps Pr: poids de matière fraîche P 5 : poids de matière sèche
l
Chapitre 1 : L 'eau et la plante
1.3.2.2. La teneur relative en eau ou contenu relatif en eau (RWC):
C'est le rapport de la masse d'eau présente dans l'échantillon à un moment donné et la masse d'eau maximale des cellules en pleine turgescence (Deparceveau et Buber, 2007). Un échantillon à la pleine turgescence est un échantillon dont les cellules sont à leur degré d'hydratation maximale.
Ce paramètre est très utilisé en physiologie. Il est déterminé comme suit : RWC= ((PF-Ps) I (PFTurg- Ps)).100 (Turg: turgescent)
1.3.2.3. Le potentiel hydrique 'Ph et ses composantes
Le potentiel hydrique, désigné couramment par 'l'h, est une grandeur thermodynamique intensive qui reflète le statut énergétique de l'eau dans la plante et le sol (représente l'énergie de liaison de l'eau avec le milieu) et détermine le sens des échanges d'eau entre deux points de l'ensemble formé par le sol, le végétal et l'atmosphère. Le 'l'h est négatif ou nul. Il est admis que le potentiel hydrique de l'eau libre est égal à zéro.
L'eau se déplace toujours d'une région de potentiel hydrique élevé vers une région de potentiel hydrique faible (Laval-Martin et Mazliak ,1995; Hopkins, 2003).
Le potentiel hydrique dans l'air est presque toujours le plus bas que celui du sol, ce qui explique que les plantes sont traversées par un flux d'eau ascendant. Il est exprimé en mégapascal (MPa) ou en bar (lMPa = lObar).
Pour décrire la situation osmotique des cellules végétales, on utilise la notion de potentiel de l'eau ou de la variation du potentiel hydrique. Donc la forme la plus générale du potentiel hydrique se formule de la manière suivante :
ô'I'= ô'l'0
+ô'l'n +ô'l'm.
Avec : ô 'I' = variation du potentiel de l'eau de la cellules ô '1'0
= variation du potentiel osmotique. ô 'l'n = variation du potentiel de turgescence.
Chapitre 1: L'eau et la plante
1.4. L'absorption de l'eau par les plantes
L'eau est absorbée ou rejetée, sous forme liquide ou de vapeur, par les végétaux aquatiques et les végétaux terrestres qu'ils soient pourvus ou non de racines. Si l'appareil racinaire n'existe pas, toute la surface de l'individu assure l'absorption, c'est le cas de plantes aquatiques (Mousses, Solvinacée, Hydroptèridée ), de spermaphytes, mais aussi de toutes les bryophytes aériennes qui s'imbibent d'eau de pluie, de rosée et de vapeur d'eau (Gorenflot,
1998).
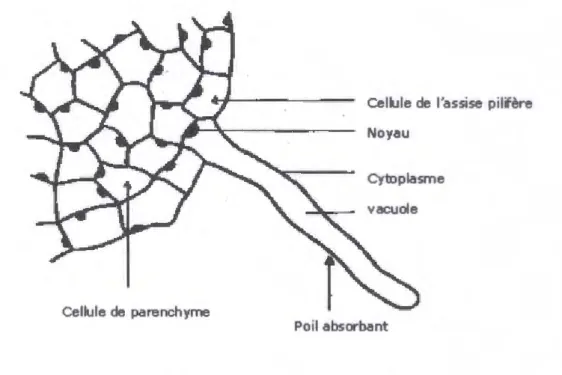
Chez les plantes terrestres, la plus grande partie de l'eau prélevée dans le sol par la plante y pénètre par les parties les plus jeunes des racines (les parties non subérifiées) qui sont les poiles absorbants (figure n°2). Les poiles absorbants, situés à plusieurs millimètres de
l'extrême des racines, représentent une énorme surface absorbante à partir des poils absorbants
(Raven et al., 2000). Ce sont des cellules géantes, très allongées, leur densité est considérable Gusqu'à 2 500 par cm2 chez les graminées) ; leur nombre peut atteindre, pour une plante de moyenne importance, plusieurs milliards. Au total pour un pied de seigle on a pu évaluer la longueur totale des racines à 250 km et leur surface absorbante à 470 m2.
Ces poils absorbants constituent les organes de prélèvement des ions minéraux dans la
solution du sol à partir des complexes colloïdaux (Laval-Martin et Mazliak, 1995) (figure n°2).
Cellule de parenchyme
Cellule de l'assise pilifère Noyau
Cytoplasme vacuole
Poil absorbant
Chapitre 1: L'eau et la plante
Les poils absorbants sont rares chez les arbres (chêne, hêtre), et même absents chez beaucoup de conifères (pin) ; ils sont alors remplacés par un feutrage de champignons symbiotes (mycorhizes) (figure n°3).
Les mycorhizes sont des filaments de mycélium (appareil végétatif des champignons formé de filaments souterrains) pénétrant dans la partie superficielle de la racine d'une plante. Leur structure est donc une association entre des cellules de champignon et celles de la racine de la plante. On a ici à faire à une relation de symbiose (à bénéfices réciproques) entre le végétal et le champignon. Expérimentalement, on constate que les arbres mycorhizés se développent beaucoup mieux, surtout lorsque le sol est pauvre en ions minéraux, et résistent mieux à la sècheresse (figure n°3).
Mycorhyze
Figure n°3: Mycorhizes ( Anonyme, sd)
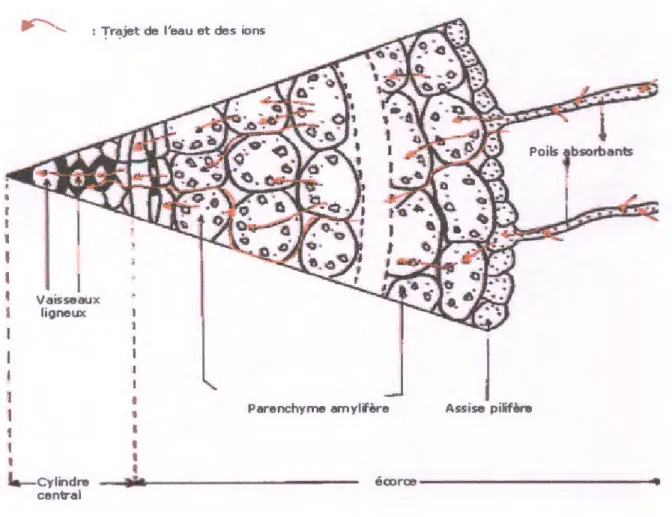
L'eau absorbée par les racines traverse le cylindre corticale, dont au moins l'assise externe peut être différenciée en exoderme et dans le cylindre conducteur, dès qu'elle arrive aux éléments conducteurs du xylème, l'eau monte à travers la racine et la tige jusqu'au feuilles, d'où la plus grande partie se perd dans l'atmosphère (Raven et al., 2000).
Chapitre I: L'eau et la plante
: "'fr~jet de l'eau et des ions
t 1
'
1 l Vaisseaux ligneux•
J.-cylindre central 1 1 1'
4Parenchyme amylifère Assise pilifère
écorce---Figure n°4: Coupe transversale d'une racine (Anonyme, sd)
L'absorption de l'eau est un processus passif due à la différence négative entre le potentiel hydrique du poil absorbant et celui du sol (Heller et al., 1998). Cette absorption est liée étroitement à la transpiration, qui crée un appel d'eau le long de la tige jusqu'aux feuilles.
L'eau se déplace du sol aux poils absorbants par un mécanisme osmotique simple, lorsque la pression osmotique des poils absorbants est supérieure à celle de la solution du sol, la perméabilité des cellules des poils absorbants augmente mais leur pression osmotique baisse à celle des cellules corticales adjacentes, de cette manière, l'eau continue à se déplacer d'une cellule à une autre jusqu'à ce qu'elle arrive aux vaisseaux du xylème.
L'eau absorbée est propulsée à l'intérieur des vaisseaux du xylème racinaire par une force qui résulte de la différence entre la pression de la solution du sol et celle de la sève brute, appelée la force de pression racinaire. Il y a également une autre force par laquelle l'eau pénètre dans les racines qui est la force d'absorption passive due à la transpiration (Aboul3ata,
Chapitre I: L'eau et la plante
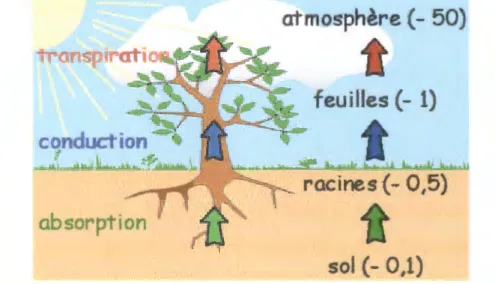
atmosphère (- 50)
feuilles (-
1)
t
absorption
sol (- 0,1)
Figure n°5: La plante et son environnement hydrique. L'eau absorbée dans le sol par les racines est conduite dans toutes les parties de la plante. Une partie est éliminée dans l'atmosphère par la transpiration. A droite, sont notées les valeurs du potentiel hydrique en Méga-Pascal (MPa) des différents partenaires (Prat et Rubinstein, 2007).
Une fois absorbée, l'eau remonte à l'intérieur de la plante. Elles passent dans les interstices entre deux cellules. Une fois arrivée en haut du végétal, l'eau s'évapore.
Le flux d'eau qui traverse la plante dépend:
•!• D'une part, de la différence de potentiel hydrique existant entre les deux pôles, sol et atmosphère;
•!• D'autre part, des différentes résistances du circuit entre ces deux pôles : résistance sol-racine, résistance dans le végétal (racines, tiges et feuilles), résistance stomatique (Minier, 2008).
Chapitre II: Le déficit hvdrique
11.1. Notion de sécheresse et de déficit hydrique
Levitt, (1980) définit la sécheresse comme une période caractérisée par un manque de
pluies qui conduit le plus souvent à un stress hydrique. Il y'a sécheresse dés lors que l'eau devient facteur limitant de la croissance et du rendement.
Selon Baldy et al. (1993), le stress dû à un manque d'eau est un déficit hydrique dans la plante qui se traduit par une perte du potentiel de production, pour les météorologues, la sécheresse est définit comme une période de 15 jours sans une pluie appréciable, en agriculture, la sécheresse est une période sèche qui conduit à une perte de rendement en
dessous de celui attendu sous des conditions optimales d'arrosage. Pour les physiologistes des plantes, la sécheresse est plus qu'un manque de précipitation: c'est l'interaction d'au moins 7 stress environnementaux (la faible disponibilité du l'eau de sol, une demande évaporativé élevée, une forte température, une forte irradiation solaire, le dessèchement du sol,
l'indisponibilité de nutriments et l'accumulation des sels dans la couche arable).
Le déficit hydrique est un phénomène courant durant le cycle de développement des plantes. Il est lié à la réduction d'humidité du sol et à l'augmentation de la demande évaporatoire.
D'un point de vue .physique le stress hydrique résulte d'un abaissement du potentiel hydrique de la plante ('V) en dessous d'une certaine valeur, dépendant du génotype et des caractéristiques du milieu, il constitue enfin le principal facteur limitant la croissance et les rendements.
Un déficit hydrique se produit lorsque l'eau disponible pour la plante ne lui permet pa~
de répondre à la demande climatique, dans se cas la perte en eau par la plante est inférieur au maximum qui serait observé en conditions hydriques non limitantes. Un déficit hydrique peu résulter d'une faible disponibilité en eau dans la portion de sol explorée par les racines et /ou d'une forte demande évaporative au niveau des feuilles (quantité d'eau transpirée est supérieur à la quantité d'eau absorbée) (Lecoeur et Guilioni, 2005).
11.2. Mécanismes d'adaptation d~s plantes au déficit hydrique
Pour lutter contre le manque d'eau, les plantes développent plusiems stratégies adaptatives qui varient en fonction de l'espèce et des conditions du milieu.
La résistance d'une plante à une contrainte hydrique peut être définie, du point de vue physiologique, par sa capacité à survivre et à s'accroître, et du point de vue agronomique, par
Chapitre II: Le déficit hvdrique
La résistance globale d'une plante à la sécheresse apparaît comme le résultat de
nombreuses modifications phénologiques, anatomiques, morphologiques, physiologiques et
biochimiques (Amor et al., 2005).
Bretaudeau et al. (1994) pensent que les mécanismes physiologiques d'adaptation à la
sécheresse sont génétiquement contrôlés
11.2.1. Esquive ou échappement
Les plantes qui peuvent esquiver la sécheresse ont la capacité de réaliser leur cycle
complet de développement avant la manifestation d'important déficit hydrique du sol
(Annerose et Comaire, 1994)
Comme les plantes de désert qui sont éphémères, autrement dit, dont la durée de vie
est courte, les éphémérophytes germent, croissent et fleurissent immédiatement après les
pluies saisonnières. Elles accomplissent donc leur cycle de développement durant une période
favorable et produisent des graines dormantes avant l'arrivée de la saison sèche. De la même .
façon, de nombreuses espèces annuelles arctiques accomplissent leurs cycles de
développement pendant le court été arctique et survivent à l'hiver sous forme de graines
(Hopkins, 2003). Leurs origines est le résultat de la domestication des e~Jces s~ages à des fins productives et se traduit généralement par un
raccourcisseme~~
~
~~aison,
de fructification et de maturation (Annerose et Comaire, 1994)
~
l{
t> \ •
\
--~\
-
•.
;?
111
11.2.2 Evitement : • ·
~J.
.
/
Se sont des mécanismes pour éviter la sécheresse, c'est-à-dire, mai'b.œtira des. niveaux élevées le potentiel hydrique de leurs tissus durant des périodes sans pluies significatives (Annerose et Corner, 1994).
L'évitement c'est le changement des caractéristiques morphologiques qui permet de
limiter la perte en eau, chez les plantes dites «économes» ou d'augmenter l'absorption de
l'eau chez les plantes dites« prodigues» (Lepoivre, 2003).
11.2.3. La tolérance :
La tolérance signifie que la plante survive à une dessiccation due à une sécheresse qui
n'endommage pas son protoplasme et qu'il conserve la capacité de reprendre une croissance normale lorsque le protoplasme sera réhydraté (Hopkins, 2003)
D'après Elhassani et Persoons (1994), la plante est résistante au déficit hydrique quand
elle est capable de maintenir son activité métabolique sous un faible potentiel d'eau jusqu'à
Chapitre II: Le déficit hvdrique
physiologique, dont le degré varie selon les espèces et le stade de croissance. Selon ces
mêmes auteurs, la tolérance à la dessiccation correspond à une capacité de la membrane cytoplasmique de retenir les électrolytes, donc de conserver son intégrité en cas de dessiccation .
. L'ajustement osmotique est considéré comme un mécanisme de tolérance par
.
certains.
auteurs etcomme un mécanisme d'évitement par d'autres (Ben Naceur, 1994).
L'ajustement osmotique est un mécanisme clé dans les relations plante - sécheresse, plante -salinité (Le poivre, 2003). Elle se rapporte spécifiquement à une augmentation nette de la
concentration de solutés dans les différents compartiments cellulaires tels que les sucres solubles, la proline ... , provoquée par des processus métaboliques déclenchés par le stress .
L'ajustement osmotique fait diminuer le potentiel hydrique qui devient donc plus négatif, permettant ainsi de maintenir le mouvement d'eau vers les feuilles et par conséquent leur turgescence (Hopkins, 2003).
Les paramètres d'adaptation au déficit hydrique sont rassemblés dans le tableau n° 3
Tableau n°3: Paramètre d'adaptation à la sécheresse
Paramètres Mécanismes Exemples
d'adaptation
Paramètres Esquive à la sécheresse Précocité phénologiques
Paramètres Amélioration de -Longueur et densité des racines
morphologiques l'absorption de l'eau -Profondeur des racines
-Rapport: parties souterraine/aérienne Réduction des pertes -Enroulement des feuilles
d'eau -Surface foliaire
-Port et orientation des feuilles Paramètres Capacité de -Hauteur de la plante
physiologiques remobiliser les -Longueur du col assimilas stockés
Maintien de l'activité -Teneur en chlorophylle photosynthétique
Capacité d'ajustement -Réduction de la transpiration par
osmotique fermeture des stomates
-Maintien d'un potentiel hydrique élevé
-Teneur relative en eau
-Osmorégulation (accumulation d'ions
Chaoitre II : Le déficit hvdrique
11.2.4. Acclimatation
L'acclimatation c'est un autre terme de réponse des plantes à la sécheresse, d'après Hopkins (2003) elle se rapporte à des modifications physiologiques non héritables, qui interviennent au cours de la vie d'un individu. La capacité de s'acclimater est bien sur un caractère génétique, mais les modifications produites en réponse au stress ne sont pas transm!ses à la génération suivan~. Le processus d'acclimatation à un stress est appelé résistance et les plantes qui se sont acclimatées à un stress sont dites résistantes par contre l'adaptation se rapport à de·s modifications de structure ou de fonction héritable qui augmente l'adéquation de l'organisme dans
lin
environnement stressant.11.3. Effets du déficit hydrique sur la plante
Tout les processus de la plante sont affectés par un déficit hydrique que se soit le métabolisme, l'organogenèse (production d'organe par le méristème) ou la morphogenèse (Dorès et al., 2006). Plus de ça, si une sécheresse importante survient, leurs résultats peuvent être tout simplement catastrophique (Bouttoud, 1995).
La perception de la plante à un déficit hydrique est différente, il y a une contrainte hydrique lorsque le fonctionnement de la plante est affecté sans bais~e de sa teneur en eau, et un stress hydrique lorsque il y a une baisse de la teneur en eau et qu'il affecte tous le métabolisme de la plante (Lecoeur et Guilioni , 2005).
, Le stress perçu par une plante, dépend de la résistance de l'organisme à un type de stress appliqué, avec une certaine intensité. En plus du type de stress et de son intensité, il faut également considérer la durée d'exposition. En effet, si l'intensité d'un stress est trop faible pour provoque~
des dommages irréversibles à court terme, à long terme, ce stress peut provoquer des changements plastiques, voire la mort de l'organisme (Levit, 1980).
11.3.2. Effets sur les paramètres morphologiques :
L'un des premiers effets provoqués par le déficit hydrique est une réduction de la croissance végétative. La croissance de la partie aérienne, et surtout celle des feuilles, est généralement plus sen$ible que celle des racines (Hopkins, 2003).
Ceci s'explique par le fait que la partie aérienne est plus exposée aux effets de la déshydratation de l'atmosphère, en effet, Sharp et Da vies (1989) in Bouamama,( 2006) observent un enracinement plus profond dans un sol sous contrainte hydrique que dans un sol régulièrement irrigué.
,. Selon Khaldoun et al., (1990), les aspects les plus étudiés de l'effet d'un manque d'eau sur le comportement morphologiques des plantes ont concerné essentiellement le développement
Chapitre II : Le déficit hvdrique
du système racinaire, le responsable d'une meilleure alimentation hydrique malgré une réduction en eau disponible du milieu et les modifications de la partie aérienne permettant une
amélioration de l'efficience de l'utilisation de cette eau.
11.3.2.1. Effets sur la partie aérienne :
Le déficit hydrique peut affecter la morphologie de la plante par un certain nombre de modifications adaptatives à savoir:
• • La réduction de la surface transpirante (par la production des feuilles très petites ou
même leur absence) ((Lemée, 1978; Harouni et al., 199 5; Albouchi, 2001 et Ben
chalbi, 2004)). D'après Harouni et al. (1995), le stresse hydrique réduit la turgescence
et par conséquent le pouvoir expansif des feuilles.
• Le changement de l'allure des profils des surfaces foliaires a été signalé par Lecoeur et
Guilioni (2005) chez le gombo.
• L'enroulement des feuilles telle que chez les riz ou il existe des cellules bulliformes qui. favorisent l'enroulement des feuilles ce qui réduit la surface foliaire exposée et isole
les stomates réduisant ainsi la transpiration de 46 à 48%, de même chez Festuca
arundinancae l'augmentation de la réflectance des feuilles par changement de leur
orientation ou accroissement de leur pubescence, qui crée un microclimat à l'humidité relative élevée chez les plantes en zones sèche (Henchi, 1987).
• L'épaississement des cuticules se fait sur les deux faces (cas des Eucalyptus) ou sur une seule face (cas·du Laurier) provoquant une augmentation de la résistance à la diffusion
,,...,
de vapeur d'eau (Ficher et Turner, 1978). La différence d'épaisseur de la cuticule
engendre une transpiration 85 fois plus grande chez Impatiens noli tangere
(hydrophytes) que chez Pinus si/vestris (Sclérophytes) (Schonherr, 1996).
• Dispositif augmentant la résistance à la transpiration par une cuticule épaisse parfois
cireuse, pilosité, enfermant des stomates au dessous de la surface épidermique ou
localisation dans des invaginations de la surface du limbe en gouttière de graminées (Le Mée, 1978), ou changement de la position des feuilles (El Hasani et Persoons, 1994 ).
• Pour réduire l'évaporation, conserver l'eau et diminuer les pertes d'eau il y a des
plantes qui produisent des feuilles charnues pour stoker l'eau (Hopkins, 2003) .
.i • D'après Lecoeur et Guilioni (2005) le déficit hydrique change complètement la
Chapitre II : Le déficit hvdrique " • Ben chelbi, (2004) dans une autre étude montre qu'à la fin de la sécheresse il y a une
réduction de la croissance de la hauteur de la partie aérienne chez Acacia cyanophylla
et Acacia tothis et le rapport tige/racine diminue. Durant et al., (1995), ont attribué
cette diminution de la croissance de la partie aérienne à une inhibition de l'extension
cellulaire qui présente l'une des premières réponses à un stress hydrique. Ou à la
suppression de l'initiation et l'expansion des feuilles et par l'abscission des feuilles.
11.3.2.2. Effets sur la partie racinaire :
Lorsque le sol s'assèche, les racines sont les premieres à détecter le déficit hydrique
(Mazliak, 1998). Sa croissance dans ce cas augmente par le faible potentiel hydrique et le rapport racine/ partie feuillé augmente lorsque l'eau devient rare dans le sol (Hopkins, 2003).
. La modification de la partie racinaire est le principal moyen par lequel les plantes
peuvent accroître leur possibilité d'absorption de l'eau au cours des périodes de sécheresse
prolongée. Elle correspond à une stratégie d'adaptation à la sécheresse. Cette stratégie a été
signalée chez plusieurs espèces. En effet, d'après Roy-Macauley (1999), le déficit hydrique
provoque une augmentation de la densité racinaire et de la profondeur d'enracinement chez
quelques variétés d'arachide. Aussi, les racines d'Acacia soumises à un déficit hydrique existent
à une profondeur de 30 m mais, chez Prosopis glandlose les racines vont jusqu'à 50 m (Kramer,
1980).
Les plantes adultes de luzerne (Medicago sativa) survivant à la sécheresse ont
développé en profondeur un système racinaire qui pénètre dans la nappe phréatique, la luzerne
peut donc être assurée d'être convenablement approvisionnée en eau dans les conditions de déficit alors que les plantesTmoins profondément enracinées subiraient la sécheresse (Hopkins, 2003).
Les plantes de maquis ont des racines très longues qui s'insinuent souvent très loin dans
les miniscules fissures de la roche (Ricklefs et Mller, 2005).
Selon Bretaudeau et al. ( 1994 ), un système racinaire profond et dense . permet dans
certaines conditions une meilleure alimentation en eau de la plante. Et d'après Hasani et
Persoons (1994), les plantes à enracinement superficiel et peu dense souffrent plus de déficit
hydrique que ceux à enracinement profond.
Les résultats obtenus par Benchelbi (2004), montrent qu'il y a une évolution de la
biomasse de la partie racinaire chez les deux espèces d'Acacia (Acacia cyanophylla et Acacia totilis). L'allocation de biomasse chez Argania Spinoza (L) s'est traduit par des valeurs du rapport longueur de la partie racinaire/ . hauteur de la partie aérienne, toujours supérieur chez
•
Chapitre II: Le déficit hvdrique
Selon Sharp et Davies (1979), un déficit hydrique peut augmenter la vitesse d'allongements racinaire et une diminution du rapport partie aérienne /racinaire.
Une étude de Sayar et al. (2008), sur le blé dure, a montré que sous stress hydrique, les racines produites sont plus longues, plus fines et ramifiées, un effet compensatoire apparaît entre les .paramètres .d'en.Î'acinement, le nombre et le volume de racines sont réduits, la longueur des axes principaux parait être le paramètre le plus relié à la tolérance à la sécheresse alors que la finesse racinaire y contribue à un degré moindre.
Chez Sinapis a/ba, une lignification du système racinaie avec formation d'ébauches racinaires renflées (tubérisation), contient des organes de survie assurant aux plantes une vie · ralentie, en attendant que les conditions redeviennent favorables (Vartanian, 1973)
11.3.3. Effets du déficit hydrique sur le rendement et le développement des plantes :
L'étude de Larbi et al., (1997), sur deux variétés de blé dur (waha et vitrion) en cas de déficit hydrique, montrent qu'il y a une réduction du rendement et ces composantes (le nombre d'épis et le poids de graine). Chez le gombo le déficit hydrique en phase de boutonnaisation réduit les quatre paramètres du fruits (Sawadogo et al., 2005).
Bouzouba et Abdelhamide (2003), dans leurs études sur l'arganier: Argania Spinosa (L) Skeels montrent que la germination de ses plantes est limitée par le déficit hydrique, de plus le déficit hydrique est l'irrégularité des pluies constituent le facteur limitant essentiels pour la production agricole en zone méditerrariéenne.
Les principaux effets du déficit hydrique sur les composantes du rendement sont notés dans la figure n° 6.
11.3.4. Effet du déficit hydrique sur les paramètres du statut hydrique
11.3,4.1. Sur la teneur relative en eau
Beaucoup de travaux montrent que la teneur relative en eau baisse en cas de déficit hydrique, par exemple, la réduction de la teneur relative en eau est remarquée en phase de boutannaison chez deux écotypes gombo (Sawadogo et al., 2005)
t ,, Oukarroum (2007) dans son étude sur l'orge montre qu'il y a une diminution de la tenew: relative en eau des feuilles basales et apicales après 4 et 24h de déshydratation et la teneur relative en eau varie entre 78,3 /. pour la variété « Aitbaha » et 77, l I. pour « Immozzer »
IV 0
f
1
-
·
-RÇduçtion de la biomasse.
a taiU~ des feutUcs et
antieipation de la sèflesœnœ Ré~ron d'évilfet~ M~e la~hemact::
•
A'-'-Ortement . de6~Réduction fk Ja mille de l \~nveloppe;.
• ·~ivilè èt
ta
durée d!fimcli~ des feuilles
J .. Ralentissement d
l'
émission•
~tesR~tm ~
desWies--~
·
---...---
---t---
-9'1
l
z-
A.nets de , l'êmissîon~ j,mites
m
CM tk stress ~ Ré'ducti~ de nontbrede ~nspwépi \.. ,,11. Réduction: nombted•épis
parm2 1-·I~
:::::: ' ··1~
'~Chapitre II : Le déficit hydrique Tandis que les témoins maintenaient 97, 1-94, 1 ï. après 24h. Chez l' arganier, les teneurs relative en eau diminuent au fur et à mesure que le déficit s'accentue, les teneurs en eau les plus élevées sont notées en 75% de capacité au champ avec une valeur maximale de l'ordre de 85,82% par opposition, le contenue relatife en eau le plus faible est emegistré pour la capacité au champ la plus faible, donc pour le niveau de stress hydrique le plue sévère (10%), et qui est de l'ordre 59,05% (Bezzala, 2005).
Donc la teneur relative en eau s'abaisse selon l'intensité du déficit hydrique, cette réduction est observée aussi chez le blé par Nouri, ( 2002) et Eljaafari, (1993).
11.3.4.2. Sur le potentiel hydrique foliaire
L'étude de Kasroui et al. (2002), sur l'effet du déficit hydrique chez deux variétés d'olivier à huile (chtoui et.chemiali), montre qu'il y a une diminûf on du potentiel hydrique foliaire. Les mêmes résultats ont été remarqués en cas de déficit hydrique chez l'orge (Ibriz et al., 1993), chez Phaseolus et Vigna (Roy et Zuily-Fodil, 2007).
L'effet exercé par un potentiel hydrique faible est attribué à une perte de turgescence des cellules. Selon Wetgate et Boyer (1985 in Hopkins, 2003), lorsque le potentiel hydrique d~
feuilles de mais diminuait suite à un stress hydrique, celui du xylème diminuait également, le mouvement d'eau en direction de la feuille était sous la dépendance d'un gradient de potentiel hydrique entre le xylème et la feuille.
La diminution du potentiel hydrique dans le xylème réduirait l'amplitude du gradient entre xylème et feuille, privant celle-ci de son approvisionnement en eau, alors que suffisamment d'eau est disponible pour maintenir les feuilles turgescentes, sa quantité est insuffisante pour provoquer le grandissement cellulaire.
Les effets physiologiques du déficit hydrique sur la perte de turgescence sont montrés dans la figure n °7.
Figure n° 7: La perte de turgescence chez les cellules (Anonyme, 2007)
Chapitre II : Le déficit hvdrique contre pression exercée par la paroi est égal à la pression de turgescence. Si la paroi est
plastique, elle se déforme sous l'effet de la pression de turgescence et la cellule grandit. b: Le milieu a la même concentration que la vacuole. Il n'.Y a aucun échange d'eau. La cellule
n'exerce aucune pression sur la paroi. La pression de turgescence est nulle. C'est la
plasmolyse limite. La croissance n'est pas possible.
c : le milieu est plus concentré en que la vacuole. L'eau à tendance à sortir de la cellule. C'est la plasmolyse.
11.3.5. Effets du déficit hydrique sur les paramètres physiologiques :
11.3.5.1. Effets sur la réponse stomatique :
Les stomates sont les seuls dispositifs que l'évolution a conservés pour affronter le pouvoir évaporant du milieu aérien (figure n°8). Ils contrôlent les échanges diffusifs en sens opposé de vapeur d'eau et de C02 (figure n°9) (Laffray et Louguet, 1991).
Stomate
Figure n° 8 : Les stomates (Moneo et lglesias, 2008)
J-1
20
Ce'llulesde
garde Ouverture dstomate
NoyauChapitre II : Le déficit hvdrique
Les plantes répondent à de graves déficits hydriques en fermant leurs stomates (figure
n°10) polir réguler la perte d'eau par la transpiration des feuilles suivie à une évaporation direct
de l'eau des cellules de gardes (Hopkins, 2003), selon Lecœur et Guilioni (2005), cette
fermeture est la réponse aux signaux racinaires hormonaux et hydraulique.
Figure n°10: Fermeture stomatique et diminution de la transpiration foliaires
(Stratégie d'évitement) (Cochard, sd)
Chez l'arachide, la fe~eture complète des stomates en conditions de déficit hydrique est
plus précoce pendant la phase de. floraison confirmant ainsi la sensibilité de cette phase. La
transpiration cuticulaire est observée lorsque la fermeture des stomates des plantes est
complète (Roy-Macauley, 1999).
Cette fermeture va entraîner des modifications physiologiques, morphologiques et
phénologiques. L'entrée du C02 est également verrouillée lors de cette fermeture, entraînant une perturbation de l'activité photosynthétique. La fermeture emprisonne une bonne part de
l'énergie destinée à être dissipée par transpiration, ce qui a pour conséquence l'augmentation
de la température foliaire.
Selon Teare et Kanemasu (1972), l'ouverture et la fermeture des stomates sont contrôlées
par la turgescence de leurs cellules de garde. Cette dernière dépend de l'humidité du sol, de la
température des feuilles, de l'humidité de l'air, du rayonnement incident, du vent, de la
concentration en C02 de l'air et de la chambre sous stomatique.
D'après l'étude de Bezzala (2005), plus le niveau de stress hydrique devient sévère plus la
transpiration devient faible jusqu'à s'annuler pour des stress très élevés (10%). Par ailleurs, une
transpiration maximale est relevée au niveau des lots de 50%, 75% et 30% allant de 4,26
Chapitre II : Le déficit hvdrique
11.3.5.2. Effets sur la structure membranaire :
Le départ de l'eau provoque une augmentation de la concentration des solutés, ce qui
entraîne des conséquences sur la structure et le métabolisme de la plante, donc les membranes
et les protéines sont également affectés par la dessiccation ce qui entraîne des
dysfonctionnements métaboliques.
L'étude de Sawadogo et al. (2005) sur différents génotypes de gombo (Abelmosculentus
esculentes L), lorsque le déficit hydrique intervient pendant la boutannaisation et la floraison,
montre qu'il y a une valeur plus élevée de la perméabilité membranaire traduisant une fuite
importante des électrolytes. Cette fuite traduit une rupture probable de la structure normale des
membranes cellulaires qui peut modifier le déplacement des protéines membranaires
contribuant ainsi à une perte de sélectivité des membranes, ce qui conduit à la perturbation du
métabolisme de la plante (Hopkins, 2003).
11.3.5.3. Effets sur la photosynthèse :
La photosynthèse est particulièrement sensible au stress hydrique. L'influence du déficit
hydrique sur la photosynthèse se manifeste depuis la capture de l'énergie lumineuse jusqu'a
l'assimilation.
La limitation de l'activité photosynthétique est issue de la réduction de la fourniture de
Co2
aux
sites de carboxylation dans les chloroplastes qui est la conséquence de la fermeture desstomates (Dorés et al., 2006).
Les plantes qui ont la capacité photosynthétique intrinsèque la moins affectée par le
déficit hydrique, présentent une efficience de l'utilisation de l'eau (Photosynthèse
/transpiration) plus élevée et une plus grande capacité de survie (Araus et al., 1991)
Le Lay et Pascaline (2001) dans son étude sur les mécanismes de synthèse de la
chlorophylle en cas de déficit hydrique sur l'orge montre que l'état d'hydratation a
d'importantes conséquences sur le processus de la transformation des chlorophyllide ( chlide)
en chlorophylle.
Les paramètres liés aux échanges gazeux sont les plus sensibles au déficit hydrique que
Chapitre III: Effets du déficit hvdrique sur l'accumulation des composés organiques
Au niveau cellulaire, la tolérance des plantes au stress hydrique peut s'exprimer par
l'accumulation d'osmolites (Van Rensburget et al., 1995; Kiani,2007). La plupart de ces
osmolytes sont des comp~sants azotés et des composés glucidiques et hormonaux ce qm
traduit l'importance dè ces métabolismes sous conditions stressantes.
Cette accumulation assure, d'une part, la poursuite de l'absorption de l'eau du sol et,
d'autre part, la rétention de l'eau intracellulaire et le maintien de la turgescence et des
processus qui en dépendent, l'ouverture des stomates, la transpiration, la photosynthèse et la
croissance (Levigneron et al., 1995).
111.1. Effets du déficit hydrique sur le métabolisme protéique :
Il est bien établi que la sécheresse entraîne des perturbations profondes au niveau du
métabolisme azoté de la plante. Pour la plupart des végétaux étudiés on note, en cas d'un
déficit hydrique, une protéolyse accrue, une inhibition de la protéogénèse et une
accumulation des composes azotés solubles.
Au niveau moléculaire, un ralentissement du métabolisme protéique et une
augmentation du catabolisme des protéines cellulaires en réponse à la contrainte hydrique ont .
été observés (V asquez-Tello et al., 1990 in Clavel, 2002) sur Vigna unguiculata (niébé).
D'après une étude de Hanower et Brzozowska (1975), La baisse du potentiel osmotique
au niveau des racines provoque rapidement-en moins d'une heure après l'application du PEG-600-le
flétrissement des feuilles. Cette déshydratation est accompagnée d'une diminution du taux des acides
aminés protéiques dont la teneur globale chute de 1,37mmol par gramme de la matière sèche foliaire
dans les feuilles témoins à 0,85 m mol dans les feuilles des plantes traitées au PEG (soit une baisse de
près de 40%) et d'une augmentation considérable de la fraction des acides aminés libres.
Comme le montre le tableau 4, la teneur globale en aminoacides libres a augmenté sous l'effet
du choc, dans les feuilles des étages inférieurs et s'est accrue par un facteur dans les feuilles qes étages
supérieurs. Ces dernières sont visiblement plus affectées par le traitement que les feuilles plus âgée.
L'apparition d'une quantité importante de proline, pratiquement absente dans les feuilles témoins, et
l'augmentation sensible des amides, surtout de l'asparagine. Simultanément les acides glutamiques et
Chapitre III: Effets du déficit hydrique sur l'accumulation des composés organiques
Tableau n° 4: Influence d'un choc osmotique sur les acides aminés libres et autres composés
positifs à la ninhydrine des feuilles de cotonnier (Hanower et Brzozowska, 197 5)
Composés uMol par g de la matière sèche de tissue foliaire
Feuilles supérieures Feuilles inférieures
Témoin PEG Témoin PEG
Acide cystéique 1.06 0,51 0.99 0.59 Glutathion 0,25 0,76 0,67 1,19 Acide aspartique 2,03 0,77 1,78 1,ll Asparagine* 0,34 10,17 0,45 1,70 Threonine
+
Glutaminet 2,76 9.61 4,93 8,74 Sérine 1,80 2,13 1,52 1,70 Acide glutamique 4.63 1,18 4,70 0,96Proline trace 4,97 trace 2,82
X trace trace trace trace
Glycine 0,62 0,63 0,57 0,63
Alanine 7,16 7,74 7.06 7.43
Valine trace 2,94 trace 1,24
Méthionine trace trace trace
Isoleucine trace 1,72 trace 0,73
Leucine trace 0.9 1 trace 0,59
Tyrosine trace 0,37 trace 0.22
Phénylalanine trace 0,7 1 1,86 0,33
Ethanolamine 2,89 1.29 3.66 trace
Acide y-aminobutyrique 2,15 16,02 0,92 15,02
Ornithine 0,75 1.03 trace 1,01
Lysine trace 0,49 trace 452
Tryptophane trace trace trace trace
Histidine trace 0,27 trace trace
Arginine trace trace 29,11 trace
Total 26.44 64,22 46,53
Le déficit hydrique induit une diminution du potentiel hydrique foliaire, du contenu relatif en eau, de la teneur en eau foliaire et des teneurs en protéines foliaires totales et chloroplastiques. Plus le cultivar est sensible, plus ce phénomène est marqué, sauf dans le cas des protéines foliaires totales. En réponse à la contrainte hydrique, les activités protéolytiques augmentent d'autant plus que le cultivar est sensible à la sécheresse. Dans les fractions solubles, partiellement purifiées, il a été mis en évidence des sérines, cystéines et métalloprotéinases. Quand aux aspartyl protéinases, elles semblent particulièrement impliquées dans la réponse à la contrainte hydrique puisque celle-ci induit une stimulation de leur activité, d'autant plus foqe que la plante est sensible (Roy-Macauley et Zuily-Fodil, 2007).
Chapitre III: Effets du déficit hvdrique sur l'accumulation des composés organiques
111.1.1. La proline :
Sous l'effet d'une contrainte hydrique, l'augmentation de la concentration des acides aminés, en particulier la proline a été observée par plusieurs auteurs (Diofasi et al., 1995;
Kavi kashor et al., 2005) ·
L'accumulation de la proline à l'état libre dans le cytosol est reconnue comme étant un osmolyte impliqué dans les réajustements osmotiques (Thiery et al., 2004) .
L'accumulation de la proline induite par les stress, peut être le résultat de trois processus complémentaires : stimulation de sa synthèse (Morris et al., (1969), Boggess et al., (1976) cités par Tahri, (1998) ; inhibition de son oxydation (Stewart et al., (1977) ; Rayapati
et Stewart, (1991) cités par Tahri, (1998) et/ou altération de la biosynthèse des protéines (Stewart et al., (1991) cités par Tahri, (1998).
· D'après les résultats .obtenus par (Boule knafet et al., 2003) , la teneur en proline au niveau de la deuxième feuille chez le blé dur (variété vitrons) dans le cas normal d'irrigation est faible (3.470 µmol/mg) à l'état de neuf jours de stress dû à un déficit hydrique, il a été remarqué qu'il y a une faible augmentation dans la concentration de la proline égale 3,944
µmoVmg de la matière sèche. Mais après seize jours de stress ils ont observé que la. concentration de la proline augmente considérablement ou elle atteint la valeur de 9,108
µmoVmg de la matière sèche.
Une autre étude de Monneveux et Nemmar (1987) sur la même espèce (le blé dur) a montré que l'accumulation de la proline est très importante et peut atteindre 350 µg/mg MF.
Aussi, Benkhokha (2007) montre que la teneur en proline en capacité au champ ( 40%) est de 0.26 µmol/mg MS, elle atteint 1,21 mol/mg MS au traitement 30% CC de l'ordre 4 à 5 fois la valeur initiale à 40% CC, sous stress hydrique à 20% CC une augmentation de l'ordre
de 21 à 47 fois a été illustrée sous stress hydrique sévère (10% CC) l'accumulation de la proline est déclenchée chez tous les génotypes de l'ordre de 52 à 106 à la fin de 1; expérience, ·
au stress du (6.5 % CC) exige une augmentation maximale de la teneur en proline cette
Chapitre III: Effets du déficit hydrique sur l'accumulation des composés organiques
Chez la luzerne en cas d'un déficit hydrique l'accumulation de la proline dans ces tissus foliaires atteignent pratiquement le double de celle de témoin (757.15) contre (345.72 µg/100 .
mg de feuilles) (Hirech, 2006).
Selon Louchichi (1995) le taux et la vitesse d'accumulation de la proline diffèrent selon la variété du blé dur, des teneurs particulièrement élevées sont obtenues au stade 5 feuilles. Waha atteint une teneur de 225.30 µg/100 mg MF. Bidi 7, 209, 63 et Ascade 65, 203, 70 peuvent être considérées comme des variétés ayant développé le mécanisme biochimique d'adaptation à la contrainte hydrique.
L'accumulation de la proline sous conditions déficitaires a étè reportées par plusieurs autres chercheurs, chez le blé dur et tendre (Eljaafari, 1993, Nouri, 2002), le thé (Chakraborty et al., 2002), la fève (Bousba, 2002), la luzerne (Adjal, 2006).
Donc, la proline est souvent citée comme l' osmoticum le plus largement distribuée et ·
accumulée sous les différentes conditions environnementales (Le poivre, 2003), leur rôle d' osmoticum est lié au niveau de tolérance au stress (Kauss, 1977).
Monneveux et Namrnar (1987) ont confirmé que la dynamique d'accumulation était
indépendante du stade de développement et très étroitement liées à la pluviométrie.
Bezzala (2005), attribue l'accumulation de la proline comme étant une protéolyse membranaire, qui s'accumule suite à une perturbation du métabolisme des protéines. Dans une autre étude sur Argania spinosa, Clifford et al. (1998) montrent que la cause de
l'accumulation de la proline est due à l'accumulation des sucres solubles (hexose, glucose).
Elle pourrait, également, intervenir dans la régulation du pH cytoplasmique (Pesci et Beffagne, (1984) cités par Bajji (1998); El Jaafari, (1993) ou constituer une réserve d'azote .
et de carbone utilisée par la plante postérieurement à la période du stress (Tal et Rosenthal,
(1979) cités par Bajji, (1998) ; El Jaafari, (1993).
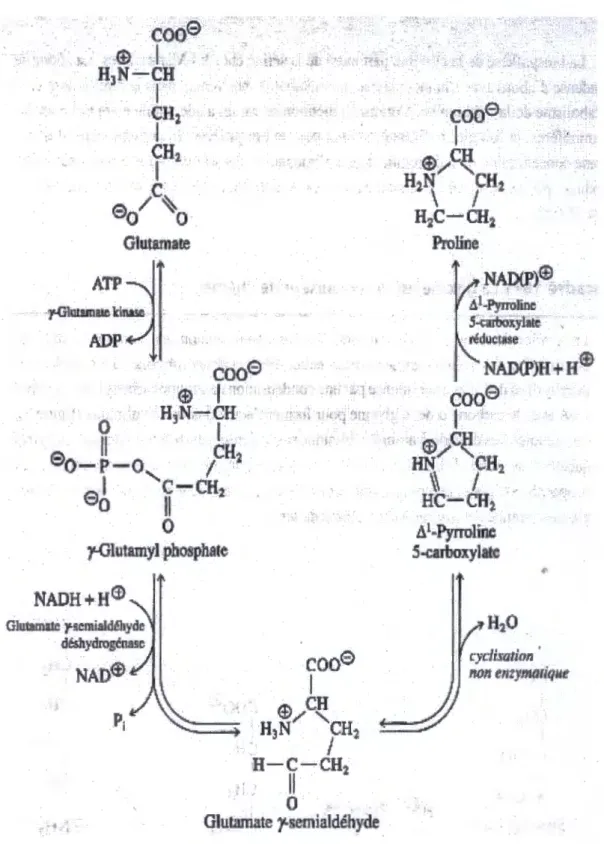
111.1.1. 1. Voies de synthèse de la proline :
La proline serait synthétisée a partir de l'acide glutamique (Hanjoh et al, 2000), une réaction qui se déroule entre le y-carboxyle du glutamate et la molécule d'ATP pour former l'acyle phosphate et donne le y-glutamyle phosphorique, acide qui se cyclera en dégageant une molécule d'H20 et forme delta pyrroline carboxylique qui ce cyclera à son tour avec une
Chapitre III: Effets du déficit hydrique sur l'accumulation des composés organiques ,.
cocJS
®
·
I
! -., · - · O H3N.-CH,Il
1
'
°'o
~-P-0
/
CR2
1
,
C-CH:2_
-90
!I
>0
y-Olutamyl phosphate
NADH+H©
' l t -(. I' l j : '·.
.
· NAD(P)@ ÂLPyrroJioe S-carboxylat1:; , ~®cr.a!!e NAD(P)H+
H®cooe
•I1
.
@/C~HN
CH2
~
I
HC-CHi
A
1-Pyrroline
5-caJboxylatcChapitre III: Effets du déficit hydrique sur l'accumulation des composés organiques•
Claussen (2005) en travaillant sur la tomate aux conditions de stress hydrique, suggère que l'accumulation de la proline serait due soit à une induction, ou activation de l'enzyme impliquées dans la biosynthèse de la proline.
Il existe deux enzymes qui commandent le métabolisme de la proline : 5 carboxilate réductase (P5CR) et proline déshydrogénase (PRODH). Le premier est responsable de sa synthèse et le deuxième de sa dégradation.
La capacité enzymatique du P5CR est faible pendant la période de l'obscurité et accélérée pendant la période de la lumière, au contraire la capacité enzymatique du PRODH est faible pendant les deux périodes ainsi l'activité de ces enzymes est relative au degré de température du milieu d'incubation, l'activité de l'enzyme PRODH fournie l'énergie nécessaire pour la fixation du nitrogène (Hanjoh et al., 2000)
111.1.1.2. Synthèse de la proline par autre voie :
La proline serait aussi synthétisée à partiJ de l'acide glutamique via l'acide 5-carboxylique 1 pyroline (p5c) mais également via l'arginine et l'ornithine (Lignowski et Slittstoesser, 1971)
Parallèlement à l'augmentation de la teneur en proline foliaire sous l' effets du stress, une baisse dans les teneurs en pigments chlorophylliens totaux (chlorophylles a et b) a été enregistrée, l'arrêt du stress permet un retour progressif aux teneurs initiales. Tahri et al.
(1997) ajoutent que la variété qui accumule plus de proline est aussi celle qui connaît la plus forte diminution de ses teneurs en pigments chlorophylliens. Dans ce cas (Leddily et al, 1993) montrent que la synthèse de la chlorophylle et de la proline sont en concurrence pour le glutamate qui est leurs précurseur commun (figure n° 12)
Chapitre III: Effets du déficit hvdrique sur l'accumulation des composés organiques
mieux caractérisées sont celles du cotonnier. Mazliak (1998) a décrit cinq classes de LEAS
-Elles sont riches en glycine et autres résidus hydrophiles (poly sérine), elles ne contiennent
pas de cystéine ou de tryptophane. Le stress hydrique entraîne la synthèse de l'EAS dans les organes végétatifs.
111.2. Effets du déficit hydrique sur le métabolisme glucidique :
Les sucres jouent un rôle prédominant dans la vie d'une plante, ils sont produits par la
photosynthèse, transportés vers les tissus profonds, canalisés vers la respiration ou convertis en composés de réserve qui. sont éventuellement dégradés en leurs constituants carbohydrates.
Le saccharose· et l'amidon sont les premiers glucides stables, issus des processus photosynthétique du cycle de Calvin et de la voie de glycolate, l'amidon s'accumule dans le chloroplaste, tandis que le saccharose synthétisé dans le cytosol est stockée dans la vacuole ou transféré vers les organes puits (Nouri, 2002).
Les sucres représentent des osmoticums beaucoup moins puissants que la proline, ils participent au maintien de la balance de la force osmotique (Bouzoubaa et al., 2002).
L'accumulation des sucres solubles est un moyen adopté par les plantes en cas de stress,
à fin de résister aux contraintes du milieu (Bajji et al., 1998).
Les sucres ont été signalés comme des osmorégulateurs potentiels. Ils jouent un rôle important dans l'ajustement osmotique, lequel est considéré comme une réponse adaptative des plantes aux conditions de déficit hydrique.
D'après les résultats de Louchichi (1995) sur le blé dur soumis à un déficit hydrique,
trouve qu'il y a une augmentation des teneurs en sucres solubles dans les fouilles de
différentes variétés, Waha accumule (1.926 mg/IOOmg MF) suivie de hedba3 (1.626 mg/lOOmg MF) et ascad 65 (1.319 mg/lOOmg/MF) bidi 7 et polinicum ont acclimulé moins·
de sucres.
Selon une autre étude chez l'arganier, l'accumulation des sucres solubles peut atteindre des valeurs de 62.63 mg/g MF et 34.23 mg/g MF, alors que le témoin accumule 7,67 mg/g MF (Bezzala, 2005) .
. D'après Hireche (20q6), le rythme d'accumulation des sucres chez la luzerne n'est pas le même que la prolirie car la teneur en sucres augmente plus sous un stress moyen que sur