Hypothyroidism in rats decreases peripheral glucose utilisation, a defect partially corrected by central leptin infusion
10
0
0
Texte intégral
Figure


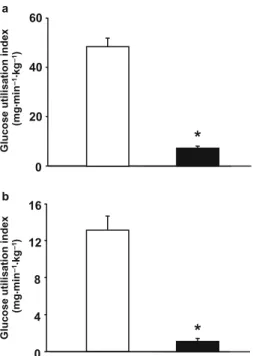
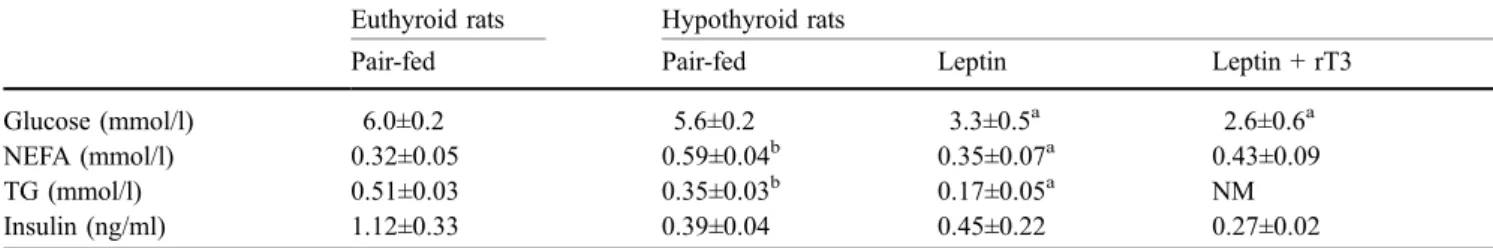
+2
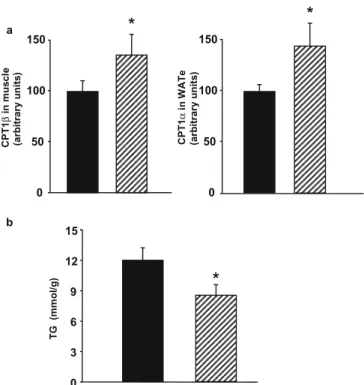
Documents relatifs