HAL Id: hal-01268084
https://hal.archives-ouvertes.fr/hal-01268084
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Impact génotoxique du CMBA, un produit de
dégradation d’un pesticide
Marie-Anne Mizon
To cite this version:
Marie-Anne Mizon. Impact génotoxique du CMBA, un produit de dégradation d’un pesticide. 2013, 20 p. �hal-01268084�
Master I Biologie et Environnement
Génétique, Ecophysiologie et Productions Végétales
IMPACT GENOTOXIQUE DU
CMBA, UN PRODUIT DE
DEGRADATION D’UN
PESTICIDE.
RAPPORT DE STAGE PRESENTE PAR :
MIZON Marie-Anne
Soutenu le 2 Septembre 2013
MEMBRES DU JURY
Aurélia Boulaflous
Mélanie Decourteix
Philippe Label
UMR 547-UBP/INRA
Physique et Physiologie Intégratives de l’Arbre
Fruitier et Forestier
Université Blaise Pascal, Campus universitaire des Cézeaux,
24, avenue des Landais, 63177 Aubière cedex, France
Responsable du stage:
Eric GOUJON
RESUME
Permettant la protection et le maintien du rendement des cultures, les produits phytosanitaires, dont les herbicides, sont, de nos jours, régulièrement utilisés dans l'agriculture. L'utilisation généralisée de ces produits chimiques ainsi que l'augmentation de la pollution dans les écosystèmes, en raison du développement industriel, peuvent provoquer une contamination de l'environnement. L'évaluation de la toxicité de ces produits chimiques ainsi que de leurs métabolites est donc devenue indispensable.
Le modèle Allium cepa a été utilisé pour étudier le potentiel génotoxique du CMBA, un des produits de dégradation de la sulcotrione. Pour cela, diverses concentrations en CMBA on été testées, et plusieurs critères ont été analysés tels que l’élongation racinaire, l’index mitotique et la fréquence d’aberrations chromosomiques. D’autre part, une mise au point de protocole sur le test des comètes a également été effectuée afin de pouvoir détecter les éventuelles lésions, simples ou double brins de l’ADN, dû au CMBA.
La réduction de la croissance racinaire ainsi qu’une augmentation de la fréquence d’anomalies chromosomiques semblent être des résultats pertinents laissant supposer que le CMBA induit un effet, aussi bien cytotoxique que génotoxique. Dans une perspective environnementale, l’étude du CMBA devient une préoccupation croissante pour la santé humaine.
Mots-clés: Allium cepa, CMBA, effet génotoxique, aberrations chromosomiques, index mitotique, élongation racinaire, test des comètes.
ABSTRACT
Pesticides, including herbicides, are regularly used in agriculture for the protection of crop yields. The use of these chemicals and the increasing pollution in ecosystems due to industrial development can cause environmental contamination. Evaluation of the toxicity of these chemicals and their metabolites has become essential these days.
Allium cepa model was used to investigate the genotoxic potential of CMBA, a degradation product of
sulcotrione. Various concentrations of CMBA have been tested on several criteria such as root elongation, mitotic index and chromosomal aberrations frequencies. On the other hand, development of the comet assay protocol was also performed in order to detect any damage, on single or double strands of DNA, due to the CMBA.
The reduction in root growth and an increase of chromosomal abnormalities appear to be relevant results suggesting that the CMBA induced effect, cytotoxic as well as genotoxic. From an environmental perspective, the CMBA study becomes a growing concern for human health.
Key words: Allium cepa, CMBA, genotoxic, chromosomal aberrations, mitotic index, root elongation, comet assay.
LISTE DES ABREVIATIONS
°C : Degré Celsius
ACP : Analyse en Composante Principale ADN : Acide DésoxyriboNucléique cm : Centimètres
CMBA : 2-Chloro-4-MesylBenzoicAcid
EDTA : Acide Ethylène Diamine Tétraacétique EMS : Ethyl MethaneSulfonate
Fig : Figure g : Gramme(s) h : Heure(s)
HSD : Honestly Significant Difference
HPPD : HydroxyPhenylPyruvate Dioxygenase LMPA : Low Melting Point Agarose
mM : milliMolaire min : minute(s) M : Molaire(s) nm : Nanomètre
NMP : Normal Melting Point PBS : Phosphate Buffered Saline TAE : Tris Acétate EDTA µL : MicroLitre
UPLC : Ultra Performance Liquid Chromatography UV : UltraViolet
SOMMAIRE
INTRODUCTION ... 1 SYNTHESE BIBLIOGRAPHIQUE ... 2 MATERIELS ET METHODES ... 4 Matériel végétal ... 4 Traitements et prélèvements ... 4 Traitement chimique ... 5 Méthodes ... 5 Elongation racinaire ... 5Fixation et coloration des extrémités racinaires ... 5
Index mitotique / Fréquence d’anomalies ... 5
Chromatographie en phase liquide en haute pression ... 6
Test des comètes ... 6
Statistiques ... 8
Résultats / Discussion ... 9
Elongation racinaire ... 9
Index mitotique ... 10
Fréquence d’anomalies ... 11
Chromatographie en phase liquide en haute pression ... 13
Test des comètes ... 14
CONCLUSION-PERSPECTIVES ... 16
REFLEXION PERSONNELLE ... 17
REFERENCES BIBLIOGRAPHIQUES ... 18
INTRODUCTION
En facilitant l’exploitation agricole et en maximisant la production, les pesticides sont, de nos jours, largement utilisés dans l’agriculture moderne. La France est le troisième pays consommateur au monde et le premier utilisateur européen de produits phytosanitaires, avec environ 70 000 tonnes déversées chaque année.
Par conséquent, la question des risques pour l’homme fait l’objet de vives controverses et constitue une priorité sur le plan santé et environnement. L'impact environnemental d'un pesticide dépend de plusieurs critères tels que son degré d'exposition, sa concentration dans l’environnement ainsi que de ses caractéristiques toxicologiques (Werf 1995). Au cours du temps, les pesticides vont être transformés en métabolites, par divers processus, qui peuvent présenter une toxicité égale voir supérieure à la matière active d’origine. L’écotoxicologie est une des disciplines qui étudie les effets toxiques d’agents chimiques ou physiques sur les organismes, en comprenant les voies de transfert de ces agents, ainsi que leurs interactions avec l’environnement.
L’étude de la dégradation de la sulcotrione, un des herbicides foliaires sélectifs du maïs, est actuellement en cours au laboratoire Physique et Physiologie Intégratives de l’Arbre Fruitier et Forestier (PIAF), en collaboration avec le laboratoire de Photochimie de l’Institut de Chimie de Clermont-Ferrand (ICCF). Cette thématique est également abordée par le laboratoire Microorganismes : génome et environnement (LMGE) qui étudie le devenir des pesticides dans les sols.
Afin d’étudier les éventuels effets toxiques de ces pesticides, des tests ont été développés pour permettre l’identification d’agents causant des dommages à l’homme. Entre autre, l’utilisation de certaines plantes supérieures a permis de mettre au point des tests de génotoxicité très performants. Certaines d’entre elles présentent de nombreux avantages tels que la facilité de manipulation, un nombre réduit de chromosomes et un cycle de développement très court. Parmi elles, Allium cepa constituent un modèle fréquemment utilisé pour les études de génotoxicité.
Mon projet a donc été d’étudier l’impact génotoxique* d’un des produits de dégradation de la sulcotrione, le CMBA, par l’intermédiaire d’Allium cepa. Pour cela, l’analyse de l’élongation racinaire ainsi que de l’index mitotique a été réalisée pour déterminer la cytotoxicité* du CMBA. Par ailleurs, l’étude de la fréquence d’aberrations chromosomiques a également été effectuée pour évaluer la génotoxicité du métabolite.
SYNTHESE BIBLIOGRAPHIQUE
Les pesticides, utilisés en agriculture pour la prévention, le contrôle ou l’élimination d’organismes jugés indésirables, qu’il s’agisse de plantes, d’animaux, de champignons ou de bactéries, sont destinés à protéger les végétaux cultivables (Magdelaine, 2010) . La France est le troisième pays consommateur au monde et le premier utilisateur européen de produits phytosanitaires, avec environ 70 000 tonnes déversées chaque année. Considéré sous l’angle des quantités de substances actives utilisées, le marché français des principaux produits phytosanitaires est largement dominé par les fongicides, qui représentent 49% du tonnage employé, suivi des herbicides dont les herbicides sélectifs du maïs et des céréales, qui représentent 34% des quantités (INRA and Cemagref, 2005).
Dans le domaine de l’agriculture, les pesticides font parti des substances susceptibles d’occasionner des risques à la fois pour la santé humaine et pour l’environnement (Tron et al., 2001; Yildiz and Arikan, 2008). Il est généralement établi que l'impact environnemental d'un pesticide dépend de son degré d'exposition, qui est fonction de sa concentration dans l’environnement, ainsi que de ses caractéristiques toxicologiques (Werf, 1995). L'utilisation des herbicides, dans la lutte contre les adventices, ainsi que l'augmentation de la pollution dans les écosystèmes, en raison du développement industriel, justifie l'évaluation de la toxicité de ces produits chimiques (Yildiz and Arikan, 2008). Par conséquent, l’estimation des risques de ces herbicides est une étape clé dans la prévention de la contamination de l’environnement. En effet, d’après le rapport de l’INRA & Cemagref, 2005, il est constaté une pollution quasi générale des eaux de surface et souterraines, principalement due aux herbicides. Concernant les eaux marines côtières et les zones de transition, la connaissance de la contamination est très partielle, voire inexistante pour les substances autres que certains pesticides organochlorés*. Pour la contamination de l’air, la présence de pesticides est observée dans toutes les phases atmosphériques, en concentrations variables dans le temps et dans l’espace. Au niveau des sols, la pollution par les pesticides varie selon la typologie du sol et les produits utilisés.Parallèlement, les matières actives sont transformées en métabolites qui peuvent présenter une toxicité égale voire supérieure à la matière active d’origine (Tron et al., 2001). C’est pourquoi, leurs utilisations peuvent avoir des conséquences aussi bien toxicologiques qu’écotoxicologiques (Werf, 1995).
La toxicologie, étudiant les effets nocifs des substances chimiques sur les organismes vivants, fait appel à une multitude de connaissances scientifiques et s'intéresse à plusieurs secteurs de l'activité humaine : l'agriculture, l'alimentation, l'environnement, etc… Les espèces les plus
fréquemment utilisées sont le rat (Guruprasad et al., 2012) et le lapin (voie cutanée), où les effets recherchés sont la létalité, la baisse de la consommation de nourriture, etc… La relation dose-réponse met en corrélation la dose d'exposition et les modifications cliniques ou biologiques induites.
L’écotoxicologie, quant à elle, est située à l’interface entre l’écologie et la toxicité. Elle étudie les effets toxiques d’agents chimiques ou physiques sur les organismes en comprenant les voies de transfert de ces agents, ainsi que leurs interactions avec l’environnement.
Le mode d'action d'un herbicide inclut tous les phénomènes qui concourent à la destruction d'une plante sensible. Pour plusieurs raisons, l'accent a été mis sur les herbicides tricétones de post-levée de type sulcotrione et mésotrione car ces produits permettent un traitement au vu de la cible.
Ces herbicides foliaires sélectifs du maïs affectent la synthèse de la chlorophylle en inhibant l’enzyme hydroxyphenylpyruvate dioxygenase (HPPD), impliquée dans la biosynthèse de plastoquinone et tocophérol. Cela va aboutir au blanchiment, à la nécrose et à la mort des plantes traitées (Chaabane et al., 2005; Schulz et al., 1993).
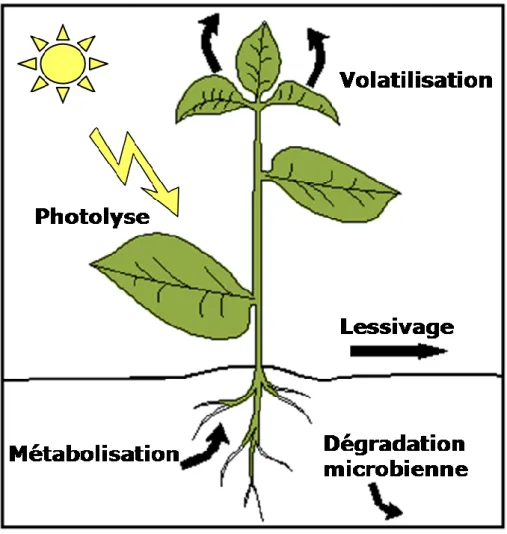
Leur dissipation peut s’effectuer selon plusieurs modes. Ils peuvent se volatiliser, être dégradés par la lumière, ruisseler ou être métabolisés (Manojlovi et al., 2013), cela dépend à la fois de facteurs biotiques et abiotiques (Fig.1) (Chaabane et al., 2005). Pour évaluer l’impact environnemental de ces herbicides, une attention particulière doit être portée sur l’importance de la présence de ces métabolites, qui peuvent varier en fonction de la pratique culturale établie sur la parcelle considérée et aussi les conditions pédoclimatiques (Mamy et al., 2005).
Afin d’étudier l’éventuel effet toxique de ces métabolites, plus de 200 tests utilisant des microorganismes, des insectes et des plantes ont été développés pour permettre l’identification d’agents causant des dommages à l’homme (Waters et al., 1988). Entre autre, l’utilisation de certaines plantes supérieures a permise de mettre au point des tests de génotoxicité très performants. Certaines d’entre elles présentent de nombreux avantages tels que la facilité de manipulation, un nombre réduit de chromosomes et un cycle de développement très court. Parmi elles, Allium cepa ainsi que Vicia faba constituent un modèle fréquemment utilisé pour les études de génotoxicité depuis les années 1930. La formation d’anomalies chromosomiques (fragments, ponts chromosomiques, micronoyaux) au sein des méristèmes racinaires révèle la capacité de cet outil à
Figure 2. Photos d’Allium cepa L. var. aggregatum seul (A) et en culture hydroponique (B).
Figure 3. Représentation schématique de l’acide 2-chloro-4-methyl sulfonyl benzoique (CMBA).
mettre en évidence les effets aneugéniques* d’agents génotoxiques (Rank and Nielsen, 1993; Sta et al., 2012).
Durant ce stage, les éventuels effets génotoxiques d’un des métabolites de la sulcotrione, l’acide 2-chloro-4-méthylsulfonylbenzoique (CMBA), ont été étudiés sur Allium cepa. D’autre part, le test des comètes est une technique récente permettant de mesurer les cassures chromosomiques. Elles sont induites par un agent génotoxique aboutissant à l'apparition de mutations chromosomiques (Yildiz et al., 2009).
Le but de cette étude est de déterminer l’effet d’un produit de la sulcotrione, le CMBA, à la fois sur la croissance racinaire, l’index mitotique, et la fréquence d’aberrations chromosomiques sur les méristèmes racinaires d’Allium cepa.
MATERIELS ET METHODES
Matériel végétal
Traitements et prélèvements
L’organisme utilisé comme test dans cette étude est une variété d’oignon nommée Allium cepa L. var. aggregatum (2n=16) (Fig. 2) appartenant à la famille des Amaryllidaceae. Cette plante bulbeuse est un modèle très largement utilisé pour évaluer le potentiel génotoxique d’agents chimiques dans l’environnement (Yildiz et al., 2009), de part son faible coût et sa culture rapide en hydroponie, permettant d’obtenir un nombre important de racines homogènes. Dans un premier temps, 21 bulbes, de diamètres équivalents, ont été choisis, rincés et placés indépendamment pour éviter toute contamination, à l’obscurité et à température ambiante, dans un tube contenant de l’eau distillée, de manière à ce que le plateau touche la surface de l’eau (Fig.2). Après 48h, les racines ayant atteintes 1 ou 2 cm de longueur, les bulbes ont été traités avec différentes concentrations de CMBA diluées dans du tampon phosphate 25 mM pour obtenir une gamme de dilution : (1.10-5M, 5.10-6M, 1.10-6M, 5.10-6M, 1.10-7M, 5.10-7M et 1.10-8M) pendant deux jours, à raison de trois bulbes par traitements. Les contrôles ont seulement été traités avec de tampon phosphate 25 mM durant la même période et un contrôle positif a été réalisé avec de l’ethyl methanesulfonate (EMS) 100mg/l, un produit connu pour ses propriétés génotoxiques, afin de valider nos conditions d’expérimentation.
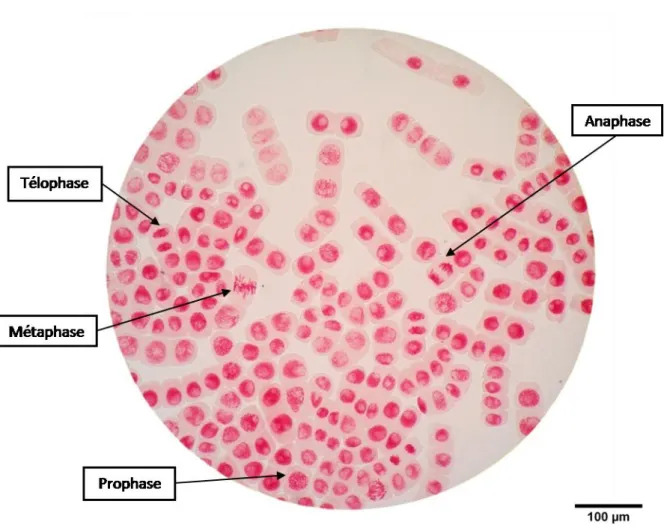
Figure 4. Les différentes phases de la mitose chez Allium cepa.
Phases de la mitose chez Allium cepa dans les cellules méristématiques des racines visualisées au microscope, à l’aide d’une coloration orcéine acétique. L’index mitotique correspond au nombre de cellules en division par rapport au nombre total de cellules, le tout ramené en pourcentage.
Nom acide 2-chloro-4-méthylsulfonylbenzoique
Molécule mère Sulcotrione
Type de pesticide Herbicide
Origine des substances Métabolisation - Photodégradation
Formule brute C14H15ClO6S
Poids moléculaire 346 g.mol-1
Densité (g.ml-1) 1,51
Dégradation DT50 au champ (jours) 8,5
Traitement chimique
La substance chimique testée est l’acide 2-chloro-4-methyl sulfonyl benzoique (CMBA) (Apollo Scientific limited) (Fig.3), un des produits de dégradation de la sulcotrione. Sa formule chimique est C14H15ClO6S et son poids moléculaire est de 346 g.mol-1. (Tableau 1)
L’éthyl méthane sulfonate (EMS) a été utilisé comme témoin durant l’étude. Sa formule chimique est C3H8O3S et a un poids moléculaire de 124,16 g.mol-1. C’est un composé chimique
génotoxique, régulièrement utilisé dans de nombreux tests mutagènes in vitro et génotoxiques comme témoin positif afin de confirmer le protocole (Rank and Nielsen, 1997; Yuet Ping et al., 2012).
Méthodes
Elongation racinaire
Pour calculer ce paramètre, deux séries de photographies d’échantillons ont été effectuées. Les bulbes mis en culture avec les divers traitements de CMBA ont été photographiés avant les traitements et 48h après. Pour chaque bulbe, une dizaine de racines ont été mesurées, à l’aide du logiciel Image J, permettant de déterminer un delta entre les deux mesures. Ce calcul a été effectué de la manière suivante :
Elongation racinaire (%) = [(longueur t48h – longueur t0h) / longueur t48h * 100]
Fixation et coloration des extrémités racinaires
Après traitements, les racines sont nettoyées à l’eau ultra-pure et les deux derniers centimètres sont prélevés et fixés une nuit à 4°C dans une solution Clarke (éthanol absolu et acide acétique glacial 3:1).
Les extrémités racinaires peuvent ensuite être conservées à long terme dans de l’éthanol 70% ou bien colorées pour une observation immédiate. Les extrémités racinaires sont rincées dans de l’eau distillée pendant 10min, hydrolysées dans HCL 1N pendant 5 minutes et à nouveau transférées dans de l’eau distillée. Ensuite, les racines sont colorées à l’orcéine acétique 1% durant une heure minimum. Seule la région correspondant à la zone méristématique sera observée après avoir écarté la coiffe et étalé les cellules.
Index mitotique / Fréquence d’anomalies
Les éventuels effets du CMBA sur les différents traitements ainsi que sur les contrôles ont été observés sous microscope Olympus CX41. Pour déterminer les effets du CMBA sur l’index mitotique, 3000 cellules ont été comptées par bulbe, à raison de 3 bulbes pour chacun des
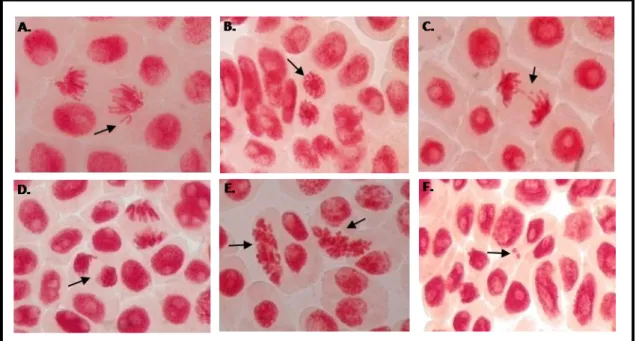
Figure 5. Exemples d’anomalies chromosomiques observées chez Allium cepa.
Anomalies chromosomiques chez Allium cepa dans les cellules méristématiques de racines, visualisées au microscope. (A.) Vagabond; (B.) Stars; (C.) Pont/Fragment; (D.) “Sticky”,
7 traitements de CMBA ainsi que les 2 témoins. Pour chaque bulbe, l’index mitotique a été déterminé sur environ 7 méristèmes racinaires en se plaçant dans 4 champs d’observation distincts, au microscope, contenant environ une centaine de cellules. L’index mitotique correspond à la proportion de cellules en cours de division en prenant en compte tous les stades de la mitose (prophase, métaphase, anaphase et télophase) (Fig. 4) par rapport au nombre total de cellules, le tout ramené en pourcentage.
Les anomalies chromosomiques sont comptabilisées au niveau des cellules en anaphase et en télophases (Rank and Nielsen, 1997). Les anomalies chromosomiques observées sont des vagabonds, des chromosomes sous forme d’étoiles (stars), des fragments, des ponts chromosomiques et des « sticky » (Fig. 5). Ces aberrations chromosomiques sont les plus fréquentes et donnent un aperçu de la génotoxicité du traitement (Panneerselvam et al., 2012). D’autres anomalies de type c-mitosis et micronoyaux (Fig. 5E, F) ont également été prises en compte.
Chromatographie en phase liquide en haute pression
Un échantillon témoin ainsi qu’un échantillon ne contenant que du CMBA de concentration 5.10-5M ont été utilisés. Les solutions qui ont servies aux traitements ont été filtrées à l’aide d’un filtre de 0.20µm déposées dans des tubes « vial » afin d’être analysées par UPLC. Le débit du solvant dans la colonne a été fixé à 0,5ml/min pour toutes les analyses et le coefficient d’absorption molaire établi à 5500l/mol/cm à pH 2,5.
Test des comètes
Ce test est basé sur la détection de cassure simple et double brin dans les noyaux des cellules. Le protocole suivi est celui décrit par Singh et al. avec quelques modifications dont la plus importante est liée au fait que ce sont des noyaux qui sont isolés et non pas des cellules (Singh et al., 1988). En effet, son protocole est basé sur des cellules animales, alors que cette étude se porte sur des cellules végétales ayant des parois.
Isolement des noyaux
De nouvelles racines d’Allium cepa ont été excisées, rincées à l’eau distillée puis délicatement découpées à l’aide d’une lame de rasoir dans 200µl de tampon (PBS + Na2EDTA
1mM). Les suspensions nucléaires seront ensuite filtrées dans un filtre 60µm réalisé en polyamide afin d’éliminer les débris cellulaires.
Figure 6 : Cellules présentant différents niveaux de dommages mis en évidence par le test des comètes. (Observations au microscope à épifluorescence).
Préparation des lames
Des lames de microscopie sont préalablement préparées avec une première couche comprenant 70µl d’agarose Normal Melting Point (NMP) 0.5% préparé dans du tampon TAE. Après une nuit, 70µl de suspension nucléaire ainsi que 150µl de LMPA 0.8% sont mélangés afin de pouvoir déposer 100µl de ce mélange sur la lame. Une lamelle est ensuite déposée sur la lame et mise 10min à 4°C pour que l’agarose se polymérise. La lamelle est ensuite délicatement retirée puis 85µl de LMP 0.5% sont ajoutés. De la même façon, les lames sont recouvertes d’une lamelle et l’ensemble sera placé 30min à 4°C.
Lyse
Ensuite, dépourvues de leurs lamelles, les lames sont aussitôt immergées dans une solution de lyse alcaline (NaCl 2.5M + Na2EDTA 1000mM + Tris 10mM, pH ajusté à 10.5 + Triton X100
1% + DMSO 10%) pendant 24h à 4°C, permettant de lyser les membranes nucléaires et de dénaturer l’ADN, c’est-à-dire séparer les deux brins de la double hélice.
Dénaturation et électrophorèse
Le lendemain de la lyse, les lames ont été rincées dans la solution d’électrophorèse (NaOH 300mM + Na2EDTA 1mM) afin d’éliminer les sels et les détergents qui pourraient empêcher la
migration de l’ADN durant l’électrophorèse (comme le NaCl). Les lames sont ensuite immergées dans la solution d’électrophorèse pendant une heure à 4°C permettant la dénaturation et le déroulement de l’ADN. Cette étape doit être réalisée en conditions alcalines (grâce au NaOH) afin de permettre la mise en évidence à la fois des coupures double brins mais aussi simple brin.
Les lames ont alors été déposées les unes à coté des autres dans une cuve d’électrophorèse horizontale. L’électrophorèse est réalisée durant 25min pour un voltage de 25V et un ampérage de 300mA. Les molécules d’ADN étant chargées négativement du fait de la présence de phosphates, migrent vers l’anode lorsqu’elles sont soumises à un champ électrique. La vitesse de migration ne dépend que de la taille des éléments puisque la concentration en agarose et le voltage sont constants. Par conséquent, les molécules d’ADN intacts vont peu migrer (du fait de leur haut poids moléculaire), et vont ainsi former une sphère compacte. Un ADN endommagé va, quant à lui, voir migrer les fragments de petite taille en dehors de la sphère et donc former une image ressemblant ainsi à une comète (Fig. 6).
Révélation de l’ADN
Cette étape est précédée d’une neutralisation (solution Tris 0,4M, pH ajusté à 7,5) car le révélateur utilisé est le DAPI, un intercalant ADN, par conséquent les deux brins d’ADN doivent
être réappariés pour que la révélation puisse se faire. Les lames ont donc été placées pendant 10min dans le tampon de neutralisation puis rincées à l’eau distillée avant d’être fixées 20min dans l’éthanol froid puis séchées à l’air libre. Les lames peuvent ainsi être stockées pour une observation ultérieure. Les lames sont réhydratées dans l’eau distillée durant 5min, puis 200µl de DAPI sont déposés. Après 20min, les lames ont été rincées à l’eau distillée puis séchées avant l’observation au microscope à fluorescence.
Examen des lames
L’examen des lames est réalisé à l’aide d’un microscope à fluorescence ZEISS Axioplan 2 (Filtre 01 d’excitation 395nm ; filtre d’émission LP397) et relié par l’intermédiaire d’une caméra CDD à un analyseur d’images.
Statistiques
Dans cette étude, les analyses statistiques de variances des données ont été réalisées sur tous les échantillons testés, à l’aide du logiciel Statistix9. Le test de Dunnett’s a été utilisé pour comparer les échantillons traités par rapport aux échantillons contrôles avec une erreur seuil alpha de 0.05. Par ailleurs, le test HSD de Tukey a été utilisé afin d’établir des groupes homogènes entre certaines données. L'Analyse en Composantes Principales (ACP), quant à elle, a été réalisée avec le logiciel JMP Pro 10.
Figure 7 : Représentation schématique d’une racine.
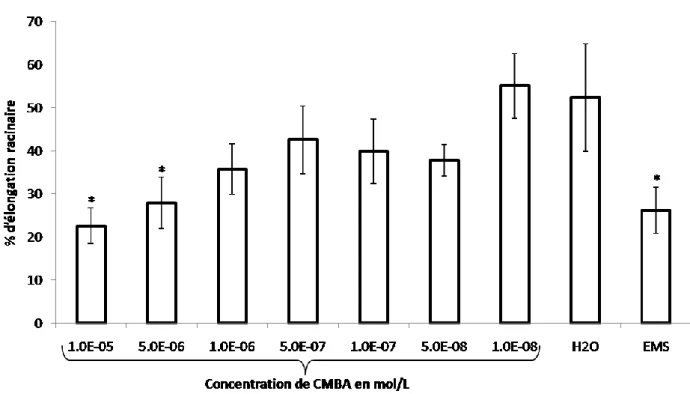
Figure 8. Mesure de l’élongation racinaire en fonction des concentrations en CMBA (mol/L).
Mesure de la croissance racinaire 48h après traitement sur des bulbes d’Allium cepa traités avec du CMBA aux concentrations : 1.10-5M, 5.10-6M, 1.10-6M, 5.10-7M, 1.10-7M, 5.10-8M, 1.10-8M, ainsi que sur des bulbes traités à l’EMS, à une concentration de 100mg/L. Ces mesures sont comparées aux mesures faites également sur les bulbes témoins (H2O). La mesure a été effectuée de la manière
suivante :
Elongation racinaire (%) = [(longueur t48h – longueur t0h) / longueur t48h * 100].
Test statistique appliqué à raison de trois répétitions par traitement : analyse de la variance suivit d’un test de Dunnett’s (test de comparaison multiple par rapport au témoin).
Résultats / Discussion
Elongation racinaire
La croissance en longueur de la racine (rhizogenèse) se produit à son extrémité, dans une région appelée apex racinaire, elle-même divisée en 3 zones : une zone de régénération de la coiffe, une zone nommée méristème racinaire où se forment les cellules de la racine et une zone quiescente où il n’y a pas de divisions cellulaires. Quand les cellules sont formées, elles s’allongent ce qui correspond à l’élongation racinaire (Fig.7)
Les mesures effectuées à partir des racines d’Allium cepa ont permises d’obtenir l’histogramme de la figure 8 qui met en relation l’élongation racinaire en fonction des diverses concentrations en CMBA. Pour le témoin négatif (H2O), l’élongation racinaire atteint 55,3%. En
revanche, les racines des bulbes mises en contact de l’EMS montrent une croissance en longueur diminuée de moitié, atteignant seulement 27.0%. Les résultats concernant les différents traitements en CMBA mettent en évidence que la concentration la plus faible (1.10-8M) atteint une élongation racinaire de 55.0%, sensiblement équivalente à celle obtenue avec le témoin négatif. De même, les concentrations les plus élevées en CMBA (1.10-5M et 5.10-6M) ont une élongation racinaire proche de celle du témoin positif. Le test statistique de Dunnett’s, quant à lui, fait ressortir le fait que l’EMS ainsi que les concentrations en CMBA de 1.10-5
M et 5.10-6M sont significativement différentes par rapport au témoin.
Les résultats mettent en évidence que les concentrations les plus élevées en CMBA ont un effet significativement différent par rapport au témoin H2O. En effet, il peut être observé un effet
dose-réponse, où les concentrations en CMBA les plus importantes induisent un retard de la croissance racinaire. Pour une concentration de 1.10-5M, le retard sur l’élongation racinaire est quasi inexistant. Cette expérience a été réalisée sur une période relativement courte. Il aurait été préférable d’effectuer les traitements en CMBA, sur une durée de 96h. La durée du traitement étant plus importante, le différentiel de taille entre le témoin négatif et les substances à analyser aurait été certainement plus élevé et cela nous aurait permis d’obtenir des mesures plus contrastées entre les différents traitements.
D’une manière générale, il est constaté une corrélation négative entre la concentration en CMBA et l’élongation racinaire, ce qui souligne un effet dose-réponse et suggère que le CMBA a un impact négatif sur la croissance racinaire. D’après Fiskesjö, le fait que la croissance racinaire diminue de plus de 45% indique la présence de substances toxiques ayant un effet sublétal sur les
Figure 9. Schéma représentant les différentes étapes de la mitose (prophase, métaphase,
anaphase et télophase).
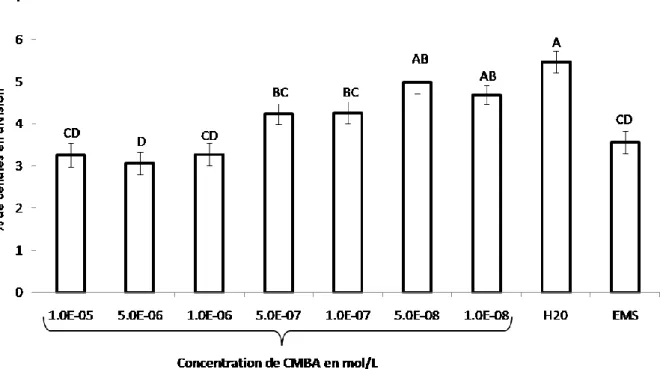
Figure 10. Détermination de l’index mitotique en fonction des concentrations en CMBA (mol/L)
Estimation de l’index mitotique 48h après traitement sur des méristèmes racinaires d’Allium cepa traités avec du CMBA aux concentrations : 1.10-5M, 5.10-6M, 1.10-6M, 5.10-7M, 1.10-7M, 5.10-8M, 1.10-8M, ainsi que sur des bulbes traités à l’EMS, à une concentration de 100mg/L. Ces mesures sont également faites sur des bulbes témoins (H2O). L’index mitotique (IM) prend en compte toutes
les phases de la mitose (prophase, métaphase, anaphase, télophase) et est calculé de la manière suivante, au niveau d’un champ d’observation:
IM (%) = [(nombre de cellules en division t48h) / nombre de cellules totales t48h * 100].
Test statistique appliqué à raison de trois répétitions par traitement : analyse de la variance suivit d’un test d’homogénéité des moyennes (test de comparaison entre moyennes).
plantes (Fiskesjö, 1985; Hidalgo et al., 1989), ce qui laisse prétendre que le CMBA a un effet cytotoxique sur Allium cepa.
Suite à ces résultats, il est montré que le CMBA affecte clairement l’élongation racinaire. Son effet peut, d’une part, altérer la maturation des cellules ou bien affecter la zone méristématique en ayant un impact sur les divisions cellulaires. Pour déterminer si le CMBA a un impact sur le processus de division cellulaire, une évaluation de l’index mitotique a été réalisée. L’index mitotique permettra de déterminer le niveau cytotoxique de l’agent chimique (Liman et al., 2010).
Index mitotique
La mitose désigne toutes les phases chromosomiques de la division cellulaire, composée de la prophase, la métaphase, l’anaphase et la télophase (Fig. 9). Elle caractérise également l’étape de séparation de chaque chromosome de la cellule mère et de leur répartition égale dans chacune des deux cellules filles. Ainsi, chaque cellule fille reçoit une copie complète du génome de l’organisme mère. Lors de l’interphase du cycle cellulaire, précédent la mitose, l’ADN alors sous forme de chromatine est répliqué, grâce à l’ADN polymérase.
L'index mitotique est l’indice qui correspond au pourcentage de cellules en mitose dans un tissu donné, ce qui permet d’estimer la fréquence des divisions cellulaires. Dans cette étude, il a été évalué à partir de comptages effectués sur des méristèmes de racines d'Allium cepa (Fig. 4).
La figure 10 représente le pourcentage de cellules en division en fonction des diverses concentrations en CMBA ainsi qu’avec l’EMS et le témoin. En ce qui concerne les racines de bulbe mises en contact avec l’eau, le pourcentage de cellules en division est estimé à 5,5%. En revanche, pour le traitement à l’EMS ce pourcentage atteint 3,5%. Ensuite, suivant les différentes concentrations en CMBA, l’index mitotique varie. En effet, les concentrations en CMBA de 1.10 -5
M, 5.10-6M et 1.10-6M ont un index mitotique avoisinant 3%. Les concentrations en CMBA de 5.10-8M et 1.10-8M ayant un pourcentage de cellules en division respectifs de 5% et 4,7% sont comparables au témoin négatif.
Par ailleurs, il a également été testé une concentration en CMBA de 5.10-5M. Cependant, à ce niveau là, quasiment aucune cellule n'était en division, seules quelques prophases étaient observables. Cela laisse supposer que pour une telle dose, le CMBA a déjà un impact tellement important qu'il n'y a plus de division cellulaire. Les étapes en aval de la mitose sont probablement bloquées ce qui concorde avec les observations faites antérieurement.
Figure 11 :Résultats de l’Analyse en Composantes Principales pour les valeurs de la fréquence d’anomalies (Fr A), de l’index mitotique (IM) et de l’élongation racinaire.
L’analyse en composante principale (ACP) a été réalisée grâce au logiciel JMP Pro10 et va permettre d’observer les corrélations existantes entre les composantes suivantes : la fréquence d’aberrations chromosomiques, l’index mitotique et l’élongation racinaire.
Le test statistique d’homogénéité effectué, démontrant la présence de groupes homogènes entre le témoin et les concentrations en CMBA les plus faibles (5.10-8M et 1.10-8M) ainsi qu'entre l'EMS et les concentrations en CMBA les plus importantes (1.10-5M, 5.10-6M, 1.10-6M), ce qui appuie les résultats décrits précédemment.
D’une manière générale, il est constaté une tendance globale qui montre que l’index mitotique diminue avec l’augmentation de la concentration du CMBA, jusqu’à devenir quasiment nul pour des concentrations égales à 5.10-5M. L’index mitotique est une mesure efficace de la cytotoxicité pour tous les organismes vivants. Le niveau cytotoxique d’un agent polluant est déterminé par la diminution de l’indice mitotique (Panneerselvam et al., 2012; Paul et al., 2013), ce qui, dans cette étude, indique que le CMBA a un effet cytotoxique sur les méristèmes racinaires d’Allium cepa en réduisant de façon importante les divisions cellulaires.
En revanche, il n'est pas possible de déterminer à quel niveau cela se produit. La réduction de l'index mitotique peut être expliquée par l’obstruction de la prophase, l'arrestation d'une ou plusieurs phases de la mitose ou le ralentissement du rythme de progression de la cellule à travers la mitose (Yuet Ping et al., 2012). Selon les travaux d’Yildiz, l’effet de certains herbicides et pesticides pourrait interférer avec le développement normal de la mitose (Yildiz and Arikan, 2008), empêchant ainsi un certain nombre de cellules d’entrer en prophase, bloquant le cycle de la mitose pendant l'interphase. Pour vérifier cela, il serait nécessaire d’effectuer des études moléculaires sur les étapes qui précédent la mitose et rechercher alors à quel stade et pour quelles raisons un checkpoint* serait bloqué.
La figure 11 met en en évidence que l'élongation racinaire et l'index mitotique sont corrélés. En effet, le méristème est constitué de cellules en division. Ces cellules, au fur et à mesure que la racine croît, vont se différencier et entrer dans la zone d'élongation. C'est pourquoi, une diminution de la fréquence de division cellulaire au sein du méristème va entraîner une diminution de la croissance racinaire ce qui explique la corrélation observée entre ces deux paramètres (Yildiz and Arikan, 2008).
Fréquence d’anomalies
Divers agents chimiques (organiques et inorganiques) et physiques (radiations ionisantes, rayonnements UV) ont la capacité d’interagir avec la molécule d’ADN et d’en modifier l’intégrité. Les dommages à l’ADN sont généralement représentés par des lésions chromosomiques induites par des agents clastogènes*. Les lésions chromosomiques sont caractérisées par l’altération du nombre (aneugénicité*) ou de la structure des chromosomes (clastogénicité).
Figure 12. Mesure de la fréquence d’aberrations chromosomiques suivant les diverses concentrations en CMBA.
Mesure de la fréquence d’anomalies chromosomiques 48h après traitement au CMBA aux concentrations : 1.10-5M, 5.10-6M, 1.10-6M, 5.10-7M, 1.10-7M, 5.10-8M, 1.10-8M. Ces études ont été réalisées sur des méristèmes racinaires d’Allium cepa au niveau des anaphases/télophases. Les fréquences ont également été déterminées sur des bulbes traités à l’EMS (100mg/L) et sur des bulbes témoins (H2O). La fréquence d’anomalies est calculée de la manière suivante:
Fréquence d’anomalies (%) = [(nombre total d’anomalies t48h) / nombre de cellules en
anaphases/télophases t48h * 100].
Test statistique appliqué à raison de trois répétitions par traitement : analyse de la variance suivit d’un test d’homogénéité des moyennes (test de comparaison entre moyennes).
La formation d’aberrations chromosomiques est évaluée chez les plantes au niveau de la zone méristématique racinaire, par un examen microscopique aux stades de l’anaphase et de la télophase (Rank and Nielsen, 1997). Ce test est destiné à détecter les agents polluants qui provoquent des anomalies touchant la structure des chromosomes, ainsi que leur potentiel génotoxique.
Les analyses cytologiques des méristèmes racinaires (Fig. 5) révèlent des changements importants dans l’organisation et la morphologie des chromosomes. Durant l’analyse microscopique, deux catégories principales d’aberrations chromosomiques, au niveau des anaphases et des télophases ont été observées: les « sticky » (Fig. 5D) et les vagabonds ou chromosomes isolés (Fig. 5A). D’autres anomalies, beaucoup moins fréquentes, de types ponts chromosomiques (Fig.
5C) et micronoyaux (Fig. 5F) ont également été observées, quelque soit le traitement appliqué.
D’apres Jiang et Marcano, les aberrations structurales sont représentées sous forme d’adhérence de chromosomes ou « sticky », de ponts chromosomiques accompagnés de rupture, de chromosomes isolés ou de micronoyaux (Jiang et al., 2000; Marcano et al., 2004). L’induction des ponts chromosomiques est une forme évidente de clastogénicité et est considérée comme l’un des tests de base de génotoxicité (Fiskesjö, 1985). Ces ponts se forment entre des chromosomes ou des fragments isolés résultant des coupures simples ou doubles brins. Les « sticky » sont définis comme étant des agglutinations de chromosomes, dues à la dégradation ou à la dépolymérisation de l’ADN, qui ont comme conséquence un aspect collant de chromosomes (Souguir et al., 2008; Yildiz and Arikan, 2008). L'induction de chromosomes isolés, quant à eux, conduit à la séparation de nombre inégal de chromosomes dans les cellules filles avec des noyaux de taille ou de forme irrégulière inégale en interphase.
Par ailleurs, d’autres anomalies de type c-mitosis (Fig. 5E) ont essentiellement été observées pour les fortes concentrations en CMBA (1.10-5M, 5.10-6M et 1.10-6M). Les c-mitosis, observées pour la première fois chez les animaux, sont une des conséquences de l’effet de la colchicine* qui entraîne l’inactivation du fuseau mitotique due à la perturbation des microtubules. Cet effet, éventuellement réversible, pourrait induire un cas d’aneuploïdie* (Fiskesjö, 1985; Liman et al., 2010). Les effets toxiques de la colchicine favorisent l’arrêt de la division cellulaire au stade de la pro-métaphase et les chromosomes continuent à se condenser, montrant ainsi une longueur réduite (Rodríguez et al., 2001).
La figure 12, quant à elle, met en évidence la fréquence d’anomalies suivant les divers traitements. Pour le témoin négatif, la fréquence d’anomalie est peu élevée (9,9%), ce qui n’est pas
le cas du témoin positif où elle atteint 16,7%. La concentration en CMBA de 1.10-5M est celle qui montre le plus fort taux d’anomalies chromosomiques (34,0%). Le résultat obtenu pour la concentration de 1.10-7M semble paradoxale par rapport aux concentrations de 1.10-6M et 5.10-7M car ces dernières n’ont pas d’effets significatifs par rapport au témoin. Dans un schéma dose-réponse classique, plus la concentration est forte, plus il y a un effet. La concentration de 1.10-7M doit donc être vérifiée si ces résultats sont reproductibles. D’autre part, le test statistique a montré que les témoins positif et négatif font parti d’un même groupe homogène, ce qui ne correspond pas aux résultats attendus. Afin d’obtenir un différentiel plus important entre ces deux témoins, il serait nécessaire d’augmenter la concentration en EMS ou utiliser un autre témoin positif tel que l’hydrazide maléique. D’une manière générale, il est constaté que le traitement au CMBA induit une augmentation du nombre d’aberrations chromosomiques par rapport au contrôle, ce qui est confirmé avec le témoin positif.
Selon Caritá et Rank, l’analyse des aberrations chromosomiques et le test des micronoyaux peuvent être considérés comme des tests efficaces pour étudier le potentiel génotoxique des traitements étudiés (Caritá and Marin-Morales, 2008; Rank and Nielsen, 1997). Dans ce cas, le CMBA aurait donc un effet génotoxique. De plus, d’après de nombreux auteurs, les anomalies dues à l’inhibition de la formation du fuseau mitotique tels que les c-mitosis ou les « sticky » reflète la haute toxicité des polluants (Lazareva, 2003; Paul et al., 2013).
Chromatographie en phase liquide en haute pression
Cette technique va nous permettre de savoir si notre produit, à savoir le CMBA, a été métabolisé par Allium cepa au cours du traitement, en observant si de nouveaux composés sont apparus. Pour cela, deux échantillons ont été analysés en UPLC : le témoin négatif (tampon phosphate) et le CMBA issu de nos traitements avec les oignons (CMBA 5.10-5M pendant 48h). Le témoin va permettre de détecter les éventuels bruits de fond dus aux déchets produits par l’oignon. Ensuite, une solution de CMBA pur (5.10-5M) sera analysée comme référence pour déterminer le temps d’élution et le profil d’absorption de cette molécule.
Figure 13. Chromatographie en phase liquide en haute pression.
Un échantillon témoin (H2O et tampon phosphate) (a), un échantillon de CMBA pur (c) d’une
concentration de 5.10-5M ainsi qu’une solution de CMBA après traitement de 48h (5.10-5M) (d) ont été analysées par UPLC. Le CMBA est quantifié, grâce à l’UPLC UV en utilisant la technique de l’étalonnage externe. Le débit du solvant dans la colonne a été fixé à 0,5ml/min pour toutes les analyses et le coefficient d’absorption molaire établi à 5500l/mol/cm à pH 2,5.
Le spectre d’absorption du CMBA (b) a également été mis en évidence au cours de cette expérience.
(a)
(d)
(c)
La figure 13 (a) met en évidence la présence d’un pic sur le profil d’UPLC qui caractérise le temps d’élution du CMBA pur, à 233nm. Chaque molécule ayant son propre spectre d’absorption, le principe de cette chromatographie est de voir si le produit seul est bien pur et si sa concentration est la même avant et après traitement. Ce pic correspond au CMBA pur et montre, grâce au profil d’absoprtion (b) qu’il absorbe à des longueurs d’ondes de 232,7nm et 282,3nm. Ce profil d’absorption correspond au même profil que celui observé à l’Institut de Chimie de Clermont-Ferrand (ICCF) (ter Halle et al., 2009). Pour l’échantillon témoin (c), aucun pic n’a été observé, seul le bruit de fond, correspondant aux déchets libérés par l’oignon, apparaît. L’échantillon correspondant à notre traitement (CMBA 5.10-5M pendant 48h) (d), a montré la présence d’un seul pic d’UPLC correspondant au temps d’élution du CMBA pur. Par conséquent, il est possible d’en déduire que le CMBA n’a pas été dégradé par Allium cepa car aucun autre métabolite n’est apparu.
Test des comètes
Ce test, basé sur la détection de cassures simples ou doubles brins dans les noyaux des cellules, est un outil efficace pour l’étude de la génotoxicité à l’échelle cellulaire. Il est applicable à tout type de tissu vivant, animal ou végétal, dont on peut isoler les cellules et extraire les noyaux. Afin de détecter l’impact génotoxique du CMBA à travers Allium cepa, nous nous sommes basés sur le protocole de Singh et al., où des modifications ont été apportées dont la plus importante est liée au fait que ce sont des cellules provenant d’un tissu végétal et non d’un tissu animal.
Auparavant, ce protocole avait été testé, au laboratoire, sur des suspensions cellulaires de l’espèce Nicotiana tabacum, mais aucun résultat concluant n’avait été obtenu car aucun noyau n’avait été observé.
Après avoir mis au point les étapes d’inclusion en gel d’agarose et choisit un système de révélation concluant, il a été constaté que l’observation au microscope n’aboutissait pas au résultat attendu. En effet, malgré les améliorations apportées, aucun noyau n’a été observé en microscopie. Par conséquent, il a été montré que l’étape visant à isoler les cellules n’était pas efficace. Après plusieurs tests sur la séparation des cellules, une autre technique consistant à isoler directement les noyaux de tissus traités a été testée. Pour cela, plusieurs méthodes visant à séparer les noyaux du reste des débris végétaux tels que des broyages ou des découpes à différents niveaux d’intensité ont été réalisées. Il s’est avéré que les découpes les plus fines permettaient un meilleur rendu sur les critères de qualité (cellules bien éclatées) et quantité (nombre de noyaux isolés interprétables). Par contre, les étapes de filtration permettant l’élimination des débris cellulaires n’a pas été concluante
puisque la plus grande partie des noyaux a elle aussi été éliminée. Cette dernière étape, déterminante pour le bon déroulement du protocole, est encore en phase de mise au point.
Une autre possibilité serait d’isoler, préalablement, les cellules du tissus végétal en retirant les parois à l’aide d’enzymes de type pectinases, afin d’obtenir des protoplastes. Ensuite, le protocole s’appuie sur une méthode classique de type Singh et al.
CONCLUSION-PERSPECTIVES
Depuis plusieurs années, les pesticides se sont installés dans le paysage agricole français, répondant à un besoin de protection des cultures contre des menaces nouvelles ou déjà existantes, dans le but de maintenir le rendement des cultures. Cependant, l’usage intensif et régulier de ces derniers ainsi que leur dégradation, donnant lieu à l’apparition de métabolites, peuvent provoquer des effets néfastes sur l’environnement. Afin de prévenir ces impacts négatifs, des études sur l’effet toxique de certains herbicides ainsi que de leurs métabolites doivent être mises en œuvre.
Le CMBA est un des principaux métabolites issu de la dégradation par oxydation de la sulcotrione. Il est notamment obtenu lors de la métabolisation de cet herbicide dans les sols, ou encore par photodégradation. Lors de ce stage, l’impact génotoxique du CMBA a été évalué de diverses manières par l’intermédiaire d’Allium cepa. L’analyse de l’élongation racinaire ainsi que de l’index mitotique a permise de mettre en évidence la cytotoxicité du CMBA, même pour une faible concentration (1.10-5M). Par ailleurs, l’étude de la fréquence d’aberrations chromosomiques, ainsi que la présence en nombre élevé de c-mitosis, souligne la génotoxicité du métabolite. Ce dernier entraîne l’inactivation du fuseau mitotique en perturbant le réseau de microtubules. Le test des comètes, dont la mise au point de protocole est en cours, sera un moyen d’appuyer le caractère génotoxique du CMBA et permettra de confirmer nos précédents résultats.
Au vu de nos résultats, il s’avère que le CMBA est toxique pour l’environnement. De même, ayant une durée de demi-vie plus importante que sa molécule mère, il devient d’autant plus néfaste pour l’environnement. Afin de valider le potentiel génotoxique du CMBA, il serait nécessaire d’effectuer des études complémentaires sur ce produit notamment sur ce qui concerne les c-mitosis. Ces anomalies atypiques apporteraient des informations sur le type de génotoxicité induite par ce métabolite. D’une manière générale, les pesticides ont un niveau de toxicité important pour l’homme avec une durée de demi-vie assez courte dans l’environnement. Par conséquent, l’impact écotoxicologique des produits de dégradation est devenu, de nos jours, une préoccupation majeure.
REFLEXION PERSONNELLE
La sauvegarde de l’environnement est un sujet d’actualité qui s’avère être de plus en plus nécessaire à prendre en compte. Pour ma part, je voulais travailler sur un sujet qui agit en faveur de l’environnement. Les domaines se rapportant à l’écophysiologie et à l’environnement entrent dans mes critères de choix pour mon travail futur.
Ce stage m’a donc permis d’avoir un premier aperçu du travail à accomplir dans un département de recherche ainsi que d’apprendre les bonnes pratiques à adopter dans un laboratoire. Mon projet au sein de l’équipe m’a permis de mettre en pratique les connaissances acquises au cours de mon cursus universitaire. De plus, au cours de ces huit semaines de stage j’ai pu me familiariser avec la démarche de la recherche scientifique et m’organiser dans mon travail afin de pouvoir mener à bien mon projet. J’ai pu acquérir de l’autonomie et de la prise d’initiative au niveau des manipulations. Enfin, ce stage m’a conforté dans l’idée que l’un de mes objectifs de carrière est de travailler pour l’environnement et le développement durable.
REFERENCES BIBLIOGRAPHIQUES
Caritá, R. and Marin-Morales, M. A. (2008). Induction of chromosome aberrations in the Allium
cepa test system caused by the exposure of seeds to industrial effluents contaminated with azo dyes. Chemosphere 72, 722–725.
Chaabane, H., Cooper, J.-F., Azouzi, L. and Coste, C.-M. (2005). Influence of soil properties on
the adsorption-desorption of sulcotrione and its hydrolysis metabolites on various soils. Journal of agricultural and food chemistry 53, 4091–4095.
Fiskesjö, G. (1985). The Allium test as a standard in environmental monitoring. Hereditas 102, 99–
112.
Guruprasad, K. P., Subramanian, A., Singh, V. J., Sharma, R. S. K., Gopinath, P. M.,
Sewram, V., Varier, P. M. and Satyamoorthy, K. (2012). Brahmarasayana protects against
Ethyl methanesulfonate or Methyl methanesulfonate induced chromosomal aberrations in mouse bone marrow cells. BMC complementary and alternative medicine 12, 113.
Hidalgo, A., Gonzalez-Reyes, J. A., Navas, P. and Garcia-Herdugo, G. (1989). Abnormal
mitosis and growth inhibition in Allium cepa roots induced by propham and chlorpropham. Cytobios 57, 7–14.
INRA and Cemagref (2005). Pesticides, agriculture et environnement.
Jiang, W., Liu, D. and Li, H. (2000). Effects of Cu2+ on root growth, cell division, and nucleolus
of Helianthus annuus L. Science of The Total Environment 256, 59–65.
Lazareva, E. (2003). Time and cell cycle dependent formation of heterogeneous tubulin arrays
induced by colchicine in Triticum aestivum root meristem. Cell Biology International 27, 633– 646.
Liman, R., Akyıl, D., Eren, Y. and Konuk, M. (2010). Testing of the mutagenicity and
genotoxicity of metolcarb by using both Ames/Salmonella and Allium test. Chemosphere 80, 1056–1061.
Magdelaine, C. (2010). Produits phytosanitaires : risques pour l’environnement et la santé. Mamy, L., Barriuso, E. and Gabrielle, B. (2005). Environmental fate of herbicides trifluralin,
metazachlor, metamitron and sulcotrione compared with that of glyphosate, a substitute broad spectrum herbicide for different glyphosate-resistant crops. Pest Management Science 61, 905– 916.
Manojlovi, D., Stankovi, D. and Rogli, G. (2013). Degradation of triketone herbicides ,
mesotrione and sulcotrione , using advanced oxidation processes. 260, 1092–1099.
Marcano, L., Carruyo, I., Campo, A. Del and Montiel, X. (2004). Cytotoxicity and mode of
action of maleic hydrazide in root tips of Allium cepa L. Environmental Research 94, 221– 226.
Panneerselvam, N., Palanikumar, L. and Gopinathan, S. (2012). Chromosomal aberrations
induced by Glycidol in Allium cepa L root meristem cells. 3, 300–304. 18
Paul, A., Nag, S. and Sinha, K. (2013). Cytological Effects of Blitox on Root Mitosis of Allium. 3,
1–7.
Rank, J. and Nielsen, M. H. (1993). A modified Allium test as a tool in the screening of the
genotoxicity of complex-mixtures. Hereditas 118, 49–53.
Rank, J. and Nielsen, M. H. (1997). Allium cepa anaphase-telophase root tip chromosome
aberration assay on N-methyl-N-nitrosourea, maleic hydrazide, sodium azide, and ethyl methanesulfonate. Mutation research 390, 121–127.
Rodríguez, E. M., Parra, M. T., Rufas, J. S. and Suja, J. a (2001). Colchicine promotes a change
in chromosome structure without loss of sister chromatid cohesion in prometaphase I-arrested bivalents. Chromosoma 110, 478–486.
Schulz, a, Ort, O., Beyer, P. and Kleinig, H. (1993). SC-0051, a
2-benzoyl-cyclohexane-1,3-dione bleaching herbicide, is a potent inhibitor of the enzyme p-hydroxyphenylpyruvate dioxygenase. FEBS letters 318, 162–166.
Singh, N. P., McCoy, M. T., Tice, R. R. and Schneider, E. L. (1988). A simple technique for
quantitation of low levels of {DNA} damage in individual cells. Experimental Cell Research
175, 184–191.
Souguir, D., Ferjani, E., Ledoigt, G. and Goupil, P. (2008). Exposure of Vicia faba and Pisum
sativum to copper-induced genotoxicity. Protoplasma 233, 203–207.
Sta, C., Ledoigt, G., Ferjani, E. and Goupil, P. (2012). Exposure of Vicia faba to sulcotrione
pesticide induced genotoxicity. Pesticide Biochemistry and Physiology 103, 9–14.
Ter Halle, A., Wiszniowski, J., Hitmi, A., Ledoigt, G., Bonnemoy, F., Bonnet, J.-L., Bohatier, J. and Richard, C. (2009). Photolysis of the herbicide sulcotrione: formation of a major
photoproduct and its toxicity evaluation. Pest management science 65, 14–18.
Tron, I., Piquet, O. and Cohuet, S. (2001). Effets chroniques des pesticides sur la santé : état
actuel des connaissances.
Waters, M. D., Stack, H. F., Brady, A. L., Lohman, P. H. M., Haroun, L. and Vainio, H.
(1988). Use of computerized data listings and activity profiles of genetic and related effects in the review of 195 compounds. Mutation Research/Genetic Toxicology 205, 295–312.
Werf, H. M. G. Van Der (1995). Evaluer l’impact des pesticides sur l'environnement.
Yildiz, M. and Arikan, E. S. (2008). Genotoxicity testing of quizalofop-P-ethyl herbicide using the
Allium cepa anaphase-telophase chromosome aberration assay. Caryologia 61,.
Yildiz, M., Ciğerci, I. H., Konuk, M., Fidan, a F. and Terzi, H. (2009). Determination of
genotoxic effects of copper sulphate and cobalt chloride in Allium cepa root cells by chromosome aberration and comet assays. Chemosphere 75, 934–938.
Yuet Ping, K., Darah, I., Yusuf, U. K., Yeng, C. and Sasidharan, S. (2012). Genotoxicity of
Euphorbia hirta: an Allium cepa assay. Molecules (Basel, Switzerland) 17, 7782–7791.
LEXIQUE
Aneugène: se dit d’un agent qui provoque des anomalies dans le nombre de
Chromosomes.
Aneuploïdie : se dit d’une cellule qui - suite à une mutation – ne possède pas le nombre normal de
chromosomes.
Checkpoint : correspond à un point de contrôle.
Clastogénécité : se dit d’un agent qui provoque des cassures de chromosomes.
Colchicine : alcaloïde tricyclique très toxique, extrait au départ des colchiques.
Cytotoxique : propriété qu'a un agent chimique ou biologique d'altérer des cellules, éventuellement
jusqu'à les détruire.
Génotoxique : Propriété d’un agent dangereux pour la santé qui exprime la capacité d’altérer de
manière directe le matériel génétique.
Organochlorés : Tout composé organique auquel on a substitué un ou plusieurs atomes
d’hydrogènes par des atomes de chlore.
RESUME
Permettant la protection et le maintien du rendement des cultures, les produits phytosanitaires, dont les herbicides, sont de nos jours régulièrement utilisés dans l'agriculture. L'utilisation généralisée de ces produits chimiques ainsi que l'augmentation de la pollution dans les écosystèmes, en raison du développement industriel, peuvent provoquer une contamination de l'environnement. L'évaluation de la toxicité de ces produits chimiques ainsi que de leurs métabolites est donc devenue indispensable.
Le modèle Allium cepa a été utilisé pour étudier le potentiel génotoxique du CMBA, un des produits de dégradation de la sulcotrione. Pour cela, diverses concentrations en CMBA on été testées, et plusieurs critères ont été analysés tels que l’élongation racinaire, l’index mitotique et la fréquence d’aberrations chromosomiques. D’autre part, une mise au point de protocole sur le test des comètes a également été effectuée afin de pouvoir détecter les éventuelles lésions, simples ou double brins de l’ADN, dû au CMBA.
La réduction de la croissance racinaire ainsi qu’une augmentation de la fréquence d’anomalies chromosomiques semblent être des résultats pertinents laissant supposer que le CMBA induit un effet, aussi bien cytotoxique que génotoxique. Dans une perspective environnementale, l’étude du CMBA devient une préoccupation croissante pour la santé humaine.
Mots-clés: Allium cepa, CMBA, effet génotoxique, aberrations chromosomiques, index mitotique, élongation racinaire, test des comètes.
ABSTRACT
Pesticides, including herbicides, are regularly used in agriculture for the protection of crop yields. The use of these chemicals and the increasing pollution in ecosystems due to industrial development can cause environmental contamination. Evaluation of the toxicity of these chemicals and their metabolites has become essential these days.
Allium cepa model was used to investigate the genotoxic potential of CMBA, a degradation product
of sulcotrione. Various concentrations of CMBA have been tested on several criteria such as root elongation, mitotic index and chromosomal aberrations frequencies. On the other hand, development of the comet assay protocol was also performed in order to detect any damage, on single or double strands of DNA, due to the CMBA.
The reduction in root growth and an increase of chromosomal abnormalities appear to be relevant results suggesting that the CMBA induced effect, cytotoxic as well as genotoxic. From an environmental perspective, the CMBA study becomes a growing concern for human health.
Key words: Allium cepa, CMBA, genotoxic, chromosomal aberrations, mitotic index, root elongation, comet assay.