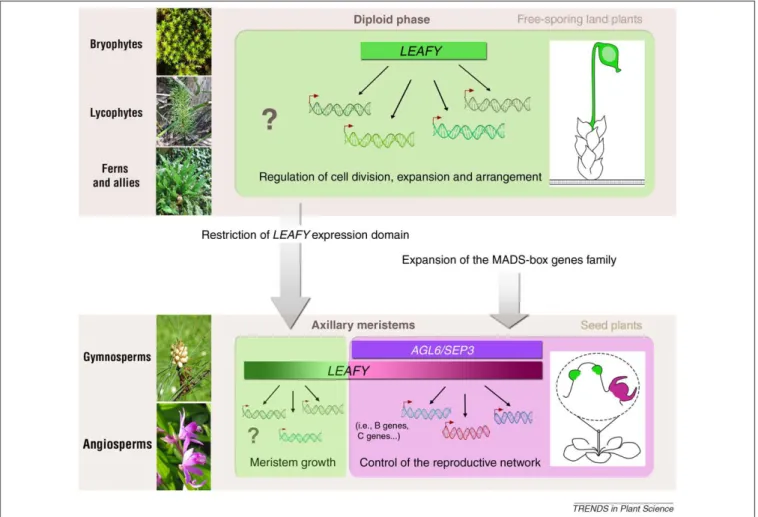
Evolution du rôle et des propriétés biochimiques de LEAFY
Texte intégral
Figure









Documents relatifs
The exchange graph sτ -tilt(Λ) is the dual graph of sτ C(Λ), i.e., the graph whose vertices are isomorphism classes of basic support τ-tilting pairs, and where two vertices are
Bien qu'il réalise que la langue des signes peut être utilisée dans l'éducation des jeunes sourds, L'Épée ne se rend pas compte que la langue des signes employée par les sourds
time scales beyond seasonal and annual cycles (Baldocchi, 2003; Saigusa et al., 2005;.. Mahecha et al. Title Page Abstract Introduction Conclusions References Tables Figures ◭ ◮ ◭
S'ils se prenaient pour des Rambo c'était leur problème, mais certainement pas le sien et elle, elle n'était pas sur terre pour se faire chier, surtout dans ce quartier de
Understanding the factors of invasion of leafy spurge in the floodplains of the Val de Saône and invasive plant interactions / natural regulators,.. in a context of
Translation repression AAAAAAAAAA RNA storage AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA Ribosome AA(n) Translation Processing g body 4E 4E 4E 4E 4E 4E 4E 4E 4G Rounds of
Two HCTs were installed on the stem and periodical readings of leaf water pressure were taken using the pressure chamber (the position of the instruments is shown in Figs 8(b) and
Substance P (SP) has been reported to produce effects on excitatory synaptic transmission in the nucleus accumbens (NAc) that are similar to those induced by cocaine.. To address
