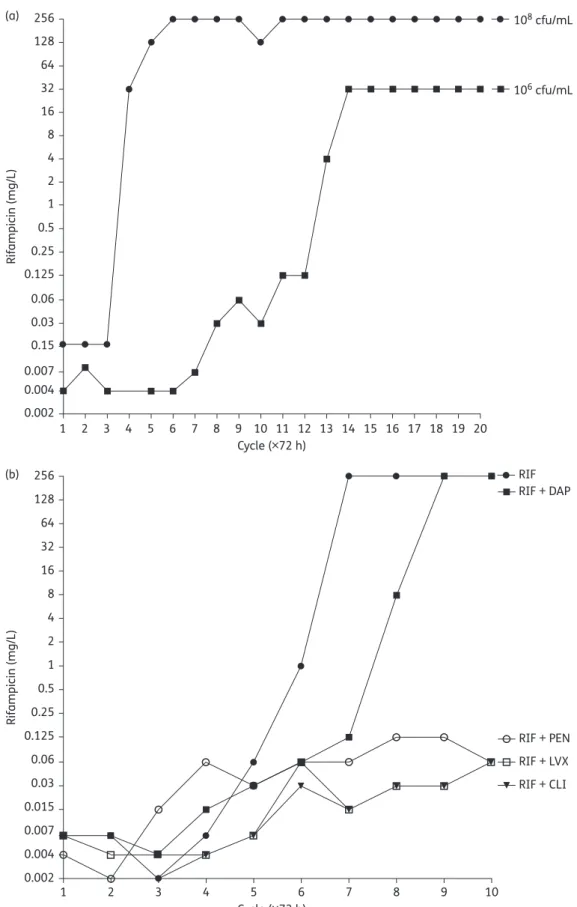
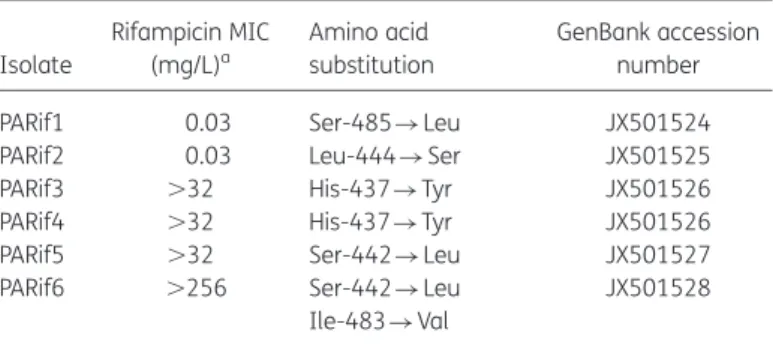
In vitro emergence of rifampicin resistance in Propionibacterium acnes and molecular characterization of mutations in the rpoB gene
Texte intégral
Figure

Documents relatifs
بقبلّلاو مسملا .شيشت رمع : ص:ص ّ خصتّلا .اهبادآو ة يّبرعلا ة غةلّلا : عرفلا .ة غةلّلا : ةرك ّ ذملا ناونع هباتك يف ي ّ بطرقلا ءاضم نبا دنع
An integrated radio-astrochronological framework of the Agrio Formation in the Andean Neuquén Basin of west-central Argentina provides new constraints on the age and the duration
Bedauerlich ist diese Unterlassung auch deshalb, da sich seit einiger Zeit eine neue Disziplin unter dem Namen Soziophysik (Helbing, 2010; Sen / Chakrabarti, 2013) zu formieren
Genes encoding NBS-LRR protein represent one of the largest and most variable gene families found in plants, with most plant genomes containing several hun- dred family
Cette technique fonctionne bien dans la généralisation d’un réseau car comme vu dans le chapitre « 2.3.2.2 - Le nombre de nœuds à utiliser par couche », moins nous avons
In order to determine if the amiR1334′s target genes (ABCB6 and ABCB20) are involved in auxin response, we generated homozygous T-DNA-insertion lines for ABCB6 and ABCB20
3 An anonymous referee has reminded me of the fact that there is also a continuous spectrum of intermedi- ate cases, parametrized by the degree to which WW information is obtained
Jung, l'anima se reflète dans la littérature dans ces figures : Dante et Ève de Milton, Béâtrice, Hélèn Troie ; sur cette base, on peut dire que toute femme dotée d'un
