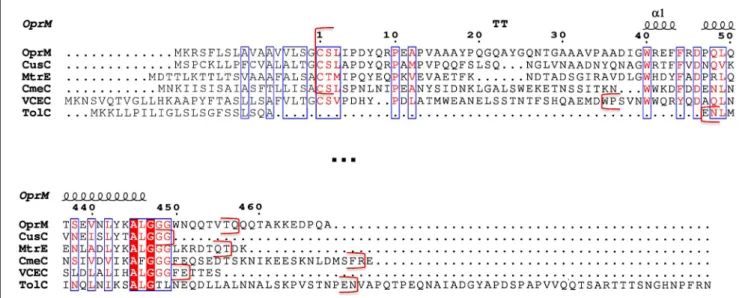
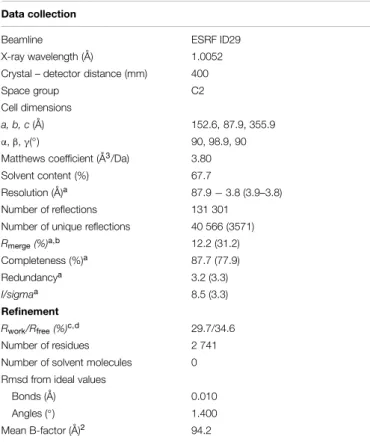
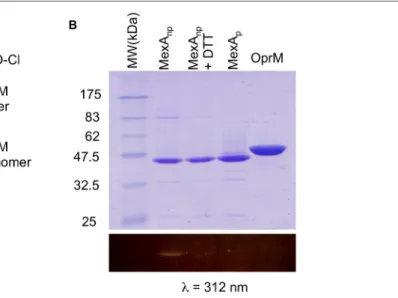
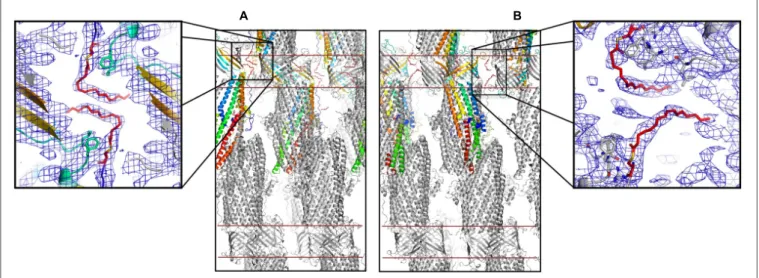
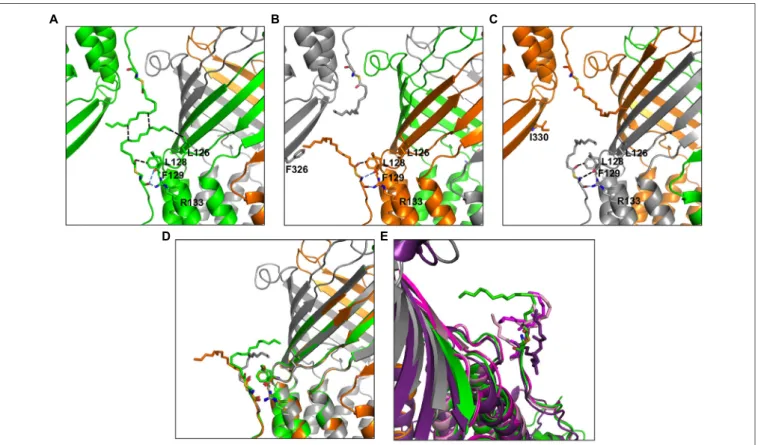
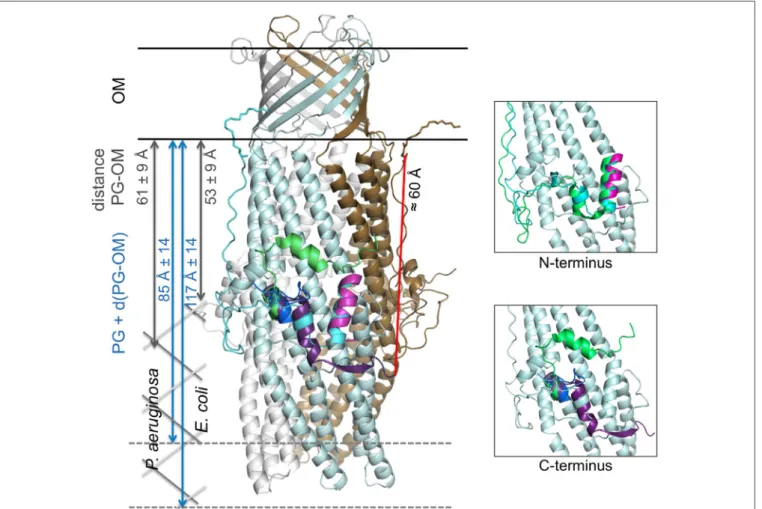
New OprM structure highlighting the nature of the N-terminal anchor
11
0
0
Texte intégral
Figure
+3
Documents relatifs