Catch-effort model used as a management tool in exploited populations: Wild boar as a case study
Texte intégral
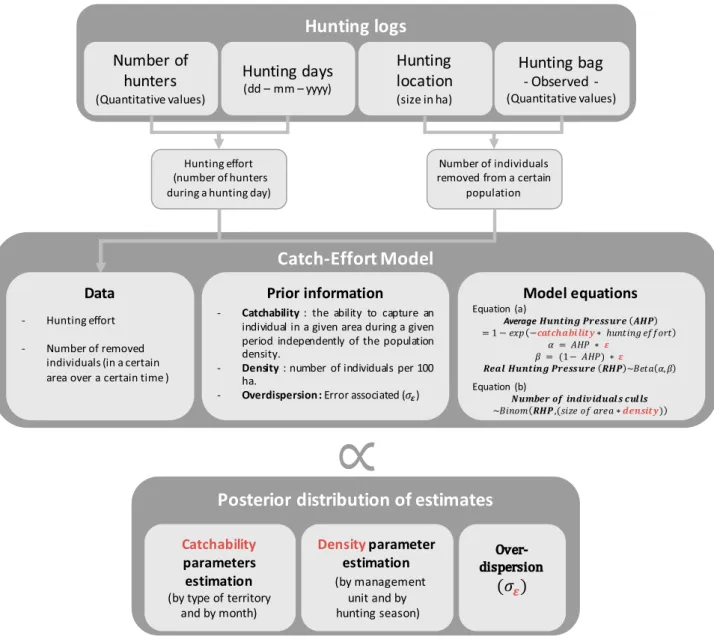
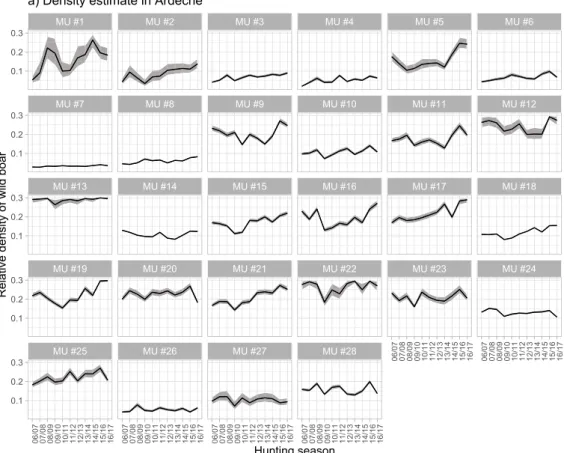
Figure

Documents relatifs
Based on the various experience of team members, this guide is organized in four main chapters: the role of numerical modeling; a tentative typology of models; a compendium of
di sviluppo, soprattutto per quelli più poveri, per- ché proprio per questi ultimi è difficile mobilitare altrove delle risorse economiche», spiega Michael Gerber, responsabile
Title: Detection of Hepatitis E virus (HEV) in a demographic managed wild boar (Sus scrofa scrofa) population in Italy Authors: Francesca Martelli, Andrea Caprioli, Martina
In the regions of southern Spain with highest cattle bTB prevalence, wildlife species such as European wild boar (Sus scrofa) and red deer (Cervus elaphus) show high prevalence
Quel que soit son statut, un agriculteur peut vendre (volontairement ou contraint par les circonstances) une partie de la production de n’importe quelle culture. Les systèmes
Application of the World Café method to discuss the efficiency of African swine fever control strategies in European wild boar (Sus scrofa) populations. Wild and feral pigs:
Account for population (control) density: analyse the estimated risk surface. but this does not account for other covariates (e.g. road-dist) Model the intesity surface using
Si par exemple on lance deux pièces de monnaie identiques, on pourrait être tenté de retenir comme ensemble fondamental = {P P, P F, F F} mais dans ce cas il n’y aurait
