HAL Id: hal-03131637
https://hal.archives-ouvertes.fr/hal-03131637
Submitted on 4 Feb 2021
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Small-scale connectivity of coralligenous habitats:
insights from a modelling approach within a
semi-opened Mediterranean bay
Vincent Rossi, Madiop Lo, Térence Legrand, Enrico Ser-Giacomi, Aurélien de
Jode, Laure Thierry de Ville d’Avray, Ivane Pairaud, Vincent Faure, Marion
Fraysse, Christel Pinazo, et al.
To cite this version:
Vincent Rossi, Madiop Lo, Térence Legrand, Enrico Ser-Giacomi, Aurélien de Jode, et al.. Small-scale connectivity of coralligenous habitats: insights from a modelling approach within a semi-opened Mediterranean bay. Vie et Milieu / Life & Environment, Observatoire Océanologique - Laboratoire Arago, In press. �hal-03131637�
Small-scale connectivity of coralligenous habitats: insights from a
modelling approach within a semi-opened Mediterranean bay
Vincent ROSSI1, Madiop LO1, Térence LEGRAND1, Enrico SER-GIACOMI2, Aurélien DE JODE3,4, Laure THIERRY DE VILLE D’AVRAY4, Ivane PAIRAUD5, Vincent FAURE1,6, MarionFRAYSSE1, Christel PINAZO1, Anne CHENUIL4
1 Mediterranean Institute of Oceanography (MIO), CNRS, Aix-Marseille University, Toulon
University, IRD, Marseille, France
2 MIT, Boston, USA.
3 Department of Marine Sciences, Tjärnö, University of Gothenburg, Strömstad, Sweden. 4 IMBE, CNRS/IRD/AMU, Endoume, Marseille, France.
5 Laboratoire d'Océanographie Physique et Spatiale (LOPS), IFREMER, Université de Bretagne
Occidentale, CNRS UMR 6523, IRD, IUEM, 29280 Plouzané, France.
6 GIPREB, Berre l’étang, France.
Keywords: habitat connectivity; larval dispersal; coralligenous assemblages; benthic organisms ; oceanic circulation ; Lagrangian modeling ; population genetics ; community composition.
Running title: Connectivity of Marseille’s bay coralligenous habitats
Abstract: Habitat connectivity is central to several key evolutionary and ecological processes, having implications for the spatial structuring of marine populations. For benthic species that have no or little mobility as adults, connectivity is evaluated by analyzing the dispersal of propagules across the seascape. We investigate fine-scale connectivity of coralligenous habitats in Marseille Bay (located in the north-western Mediterranean) sea using high-resolution cartography and a particle-tracking model ran over a range of parameters derived from the biological traits of potential target species. We present annual and seasonal means of various connectivity diagnostics measuring the retention and exchange of propagules among coralligenous patches. A synthetic view is obtained by clustering individual patches into “coralligenous provinces” (ensemble of patches tightly connected by ocean currents). When discussing our results against historical observations, we highlight some genetic breaks, sharp community changes and ecological clusters that fit well our simulated connectivity patterns. Consistent findings include reduced biodiversity along Côte Bleue, high biodiversity at Planier Island and the presence of various dispersal barriers, which evolve with dispersal durations and provide the backbone of habitat connectivity. Our results help to apprehend and test hypothesis on marine population structures, providing useful information for ecologists and conservationists.
Introduction:
The spatial structure of marine populations is a fundamental pre-requisite to understand, manage and protect marine ecosystems (Bostford et al. 2009; Dubois et al. 2016). At first order, the distribution of marine organisms is thought to be determined by the availability of habitats, which are heterogeneously distributed across the seascape, and by the movements of organisms among favorable areas. The latter process, which is referred to as “connectivity” (Rossi et al. 2016), influences key evolutionary (e.g. speciation, long-term persistence, genetic structure, local adaptation) and ecological (invasion,
colonization, demography, resilience to external perturbations) processes (Duputie & Massol, 2013). Hence, population connectivity has also been related to biodiversity (Jones et al. 2007). Finally yet importantly, habitat connectivity is a key feature to optimize when prioritizing specific areas for protection and conservation (Olds et al. 2012).
Here we evaluate the small-scale connectivity of one emblematic rocky habitat, the coralligenous, which are endemic of the Mediterranean Sea and highly fragmented at all scales (Martin et al. 2015). Our study particularly applies to benthic species (sessile and demersal): since their adult phases have no or very low motility, the inter-relationships among separated coralligenous patches are essentially controlled by the dispersal of propagules. Historical research (e.g. ecological monitoring, population genetics, artificial reefs, etc...) as well as numerous information about the physical environment (models and observations of hydrography and hydrodynamics) make the Mediterranean Sea an ideal “natural laboratory” to study fine-scale habitat connectivity. With more than 1500 species, sessile or associated (about 315 algae, 1241 invertebrates and 110 fishes, according to Ballesteros, 2006), coralligenous habitats contribute greatly to the Mediterranean biodiversity hotspot (Costantini et al. 2018), and are present along most Mediterranean shorelines (Martin et al. 2015). While some species inhabiting coralligenous habitats can be found elsewhere, there is likely more diversity per unit surface in coralligenous than in any other marine habitats as space is saturated by species and epiobiosis is frequent (Ballesteros, 2006; Cinar et al. 2020). Due to their great diversity and accessibility to coastal populations, coralligenous habitats provide ecosystem services of all types (Thierry de Ville d’Avray et
al. 2019). Species tightly depending on these habitats (e.g. red coral, spiny lobster, grouper, etc…) are
of high patrimonial and commercial values. Indeed, coralligenous landscapes are appreciated by divers and scientists for the potential of new discoveries, as many species remain understudied. Moreover, the calcareous algae forming bio-concretions, the basis of coralligenous habitats, have potential for carbon sequestration (Martin et al. 2013a). Last, the main ecosystem services provided by coralligenous (e.g. habitat, nursery, refugee, food) are crucial to sustain marine Mediterranean populations (Thierry de Ville d’Avray, 2018).
Coralligenous reefs appear particularly vulnerable in the “Anthropocene”. The coralline algae builders as well as the main animal bio-constructors have slow growth (Garrabou and Ballesteros, 2000; Torrents
et al. 2005; Rodriguez-Prieto, 2016) and narrow thermal tolerances (adapted to relatively stable
temperatures found below the thermocline). Hence, coralligenous habitats are particularly sensitive to marine heat-waves, which are likely to become stronger and more frequent with global warming (Pairaud et al. 2014). It was shown by a few historical mortality events well documented in the North-Western Mediterranean (e.g. Garrabou et al. 2009; Crisci et al. 2011) that resulted in a loss of both habitat complexity and biodiversity (Piazzi et al. 2012). As the unique limestone formation of biogenic origin (Ballesteros, 2006), coralligenous bio-constructions are also threatened by ocean acidification since calcareous algae use dissolved inorganic carbon for photosynthesis and calcification (Martin et al. 2013b). Other noticeable threats are sedimentation, nutrient enrichment (from wastewater or river outflow) and biological invasions (mainly from the algae Caulerpa cylindracea [Sonder] and
Womersleyella setacea [Hollenberg; R.E. Norris]; Piazzi et al. 2012). When a coralligenous patch is
negatively affected by mortality outbreaks, connectivity is the main process to ensure population persistence, favouring recolonization and/or access to refugee (Gerber et al. 2014).
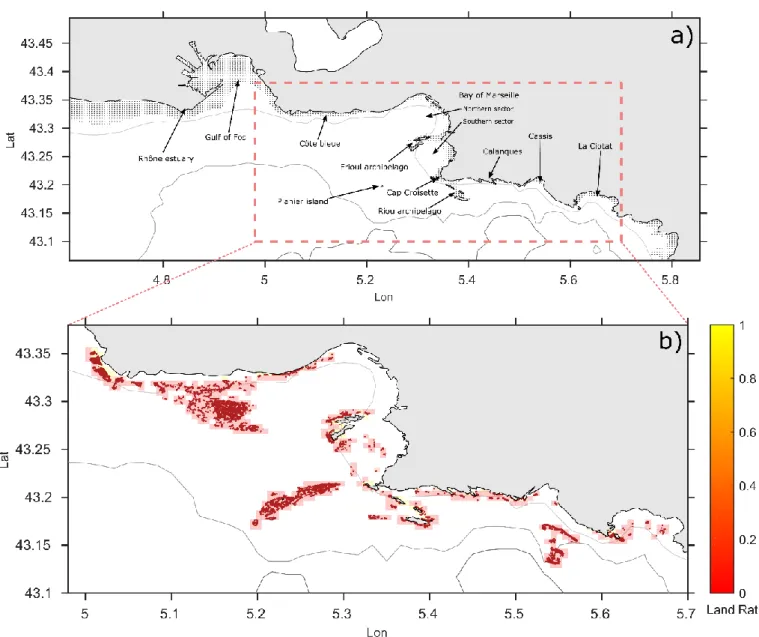
We investigate fine-scale connectivity of coralligenous habitats and its potential impacts on benthic populations focusing on the bay of Marseille, a semi-opened embayment (about 40 km by 25 km, from the Gulf of Fos to the Riou archipelago, Fig. 1a) located at the eastern entrance of the Gulf of Lion in the North-West Mediterranean Sea. The coastal circulation is influenced by various forcing conditions including complex bathymetry, strong surface winds, the Rhône river outflow and the large-scale circulation (cyclonic Northern Current NC) producing intermittent hydrodynamic patterns (Millet et al. 2018). Both dominant wind regimes, that are North-westerlies (Mistral) and South-easterlies,
respectively induce upwelling and downwelling processes (Pairaud et al. 2011; Fraysse et al. 2013). While Marseille bay can be affected by episodic arrival of eutrophic (cold, fresh and nutrient-rich) waters from the Rhône River plume (Fraysse et al. 2014), the southern sector is usually under the influence of oligotrophic waters brought by the NC (Fig. 1a). Oligotrophic conditions may also reach the northern sector on rare occasions when, instead of its usual westward offshore flow, the Northern current intrudes the continental shelf of the Gulf of Lion (Ross et al. 2016). Due to their high fragmentation (Fig. 1b) and the chaotic circulation of the bay, the connectivity of coralligenous habitat is difficult to apprehend and mostly unknown to-date.
Previous multi-specific studies of coralligenous in Marseille Bay suggested that genetic diversity differs among sites and that barriers to gene flows occur within the bay. For instance, Cahill et al. 2017 highlighted the presence of genetic differentiation for many species between the Côte Bleue and the northern bay (Fig. 1) but the presence of a physical dispersal barrier has not been investigated yet. De Jode et al. (2019) also found community differentiation between crustose algae (builder) communities located apparently in the same area, somewhere between Côte Bleue and Calanques. Only few studies already investigated the hydrodynamic connectivity of Marseille bay to compare against the genetic structures of a seaweed which is not a coralligenous dweller (Thibault et al. 2016), and to track the fate of wastewater among a few specific sites (Millet et al. 2018). An exhaustive evaluation of connectivity at small-scale and considering the precise distribution of such specific and emblematic habitats is, to our knowledge, still missing in this region.
This paper presents a statistical description of fine-scale habitat connectivity for several time scales and a range of parameters that should be relevant for some benthic species typical of coralligenous. We model the retention and exchange of propagules (e.g. eggs, larvae, spores, body fragments, rafts, etc…) driven by ocean currents among habitat patches in the bay of Marseille. We provide annually- and seasonally-averaged estimates of three complementary connectivity metrics, namely Local Retention, Self-Recruitment and Source-Sink. Furthermore, to give a synthetic view of the interlinked network of coralligenous habitats, we cluster all individual patches into a set of “provinces” (e.g. ensemble of patches, possibly disconnected geographically, which are tightly connected by ocean currents) and report mean statistics for each of them. By comparing observed spatial structure with the set of simulated spatial structures for a corresponding set of dispersal traits, one may infer the most likely dispersal traits of a target species whose dispersal abilities are unknown. We finally discuss our results against the literature, highlighting that some dispersal predictions are plausible explanations for observed genetic structures or biodiversity patterns.
Materials and Methods:
- Potential target species
Our ecosystem-based approach to dispersal consist in analyzing habitat connectivity for a range of “Pelagic Larval Durations” (PLDs; i.e. the time during which propagules drift with ocean currents) and spawning periods which are relevant for several species inhabiting coralligenous. By reviewing some information (often partial and uncertain) of their early-life traits, we intend to make a non-exhaustive list of benthic organisms to which our results could apply. The main builders of Mediterranean coralligenous habitats are calcareous red algae belonging to two families: Peyssonneliacae and Corallinaceae. In Marseille bay, species of genus Mesophyllum (Lithophyllum, respectively) are the main builders of these habitats above 20 m (below 20 m, respectively). The dispersal abilities of these calcareous red algae are thought to be very low (confirmed by population genetics results, De Jode et
al. 2019) but are mostly unknown (Norton, 1992; Opazo & Otaiza, 2007). Myriapora truncata dispersal
propagule is a lecithotrophic larva that can survive only up to 24 hours in aquarium (Ferretti et al. 2007). The gorgonian Paramuricea clavata, one of the most abundant structuring species of coralligenous habitats, also has lecithotrophic larvae suggesting low to moderate dispersal abilities, also supported by
genetic structuring at small scales (Mokhtar-Jamaï et al. 2011 and references therein). Despite the fact that supposed early-life traits would point toward high dispersal potential (e.g. Martinez-Quintana et al. 2015), other species typical of coralligenous present a clear genetic structure at small scales in the bay: the red coral Corallium rubrum (Ledoux et al. 2010, Pratlong et al. 2018) and Eunicella cavolini (Canovas Molina et al. 2018). Finally, other coralligenous species whose dispersal traits are unknown but which could have low dispersal abilities both through sexual or asexual reproduction include:
Caryophyllia inornata (Goffredo et al. 2012), sponges such Oscarella lobularis (Ereskovsky, 2010;
Ereskovsky et al. 2013). Animals, which also play a role bioconcretions like bivalves and tubercolous polychaetes, may have high dispersal abilities but no information is available to confirm this (Costantini
et al. 2018). Note that some polychaetes brood larvae in their tubes suggesting very low dispersal, if
any. Metazoan groups typical of coralligenous outcroups are bryozoans, tunicates, cnidarians and sponges whose propagules would suggest lower dispersal as compared to planktotrophic and lecithotrophic larvae.
- Cartography of coralligenous habitats
The cartography of coralligenous habitats was extracted from the dataset CARTHAM 2010-2012 by aggregating the sub-dataset #12 (Astruch et al. 2011) and #13 (Astruch et al. 2012), while some pixels were completed later by Andromède océanologie (2014). This dataset is available online on the Medtrix platform (www.plateforme.medtrix.fr, accessed in Sept. 2015) hold by the “Agence de l’Eau” and “Andromède Oceanology”. It is a fine mapping (resolution of about 20 ms) of marine habitats carried out using a lateral sonal, a multibeam sounder, including confirmation in the field by divers. High-resolution habitat maps were interpolated on our model grid and then transformed into presence/absence data, resulting in 423 discrete patches widespread across the study area (Fig. 1b).
- Hydrodynamic model
The ocean model MARS3D (Lazure & Dumas, 2008) was implemented in the RHOMA (RHOne-MArseille) configuration extending westward from the Rhône River mouth till the Cap Sicié, eastward of Marseille bay. This configuration was set up, validated and exploited by numerous previous studies (Pairaud et al. 2011; Fraysse et al. 2013; 2014; Ross et al. 2016; Millet et al. 2018). The horizontal grid mesh resolution was of 400 m with 30 vertical sigma levels. The model run used in this study covers the period of the years 2009 to 2011 with the same model setup (forcing, parametrization, and boundary conditions) described in Pairaud et al. (2011). In particular, the model was forced by the outputs from a high-resolution (3 km; 3 hours) atmospheric model allowing the simulation of short-lived wind-forced upwelling and downwelling events. To take into account the influence of the large-scale general circulation, and especially the Northern Current intrusions in the bay, the model was forced at the western and southern boundaries by the 3-hourly results of a regional hydrodynamic model under its MENOR configuration (Nicolle et al. 2009). Hourly current outputs of the RHOMA model are provided on sigma vertical levels and Arakawa-C horizontal grid. Hourly flow fields on an Arakawa-A grid at a given depth (20 m) were obtained by vertical interpolation followed by horizontal interpolation to feed in the offline dispersal model.
- Off-line dispersal model
The Lagrangian Flow Network (LFN) methodology combines network theory tools and particle-tracking model to investigate transport and dispersal induced by ocean currents. As most off-line Lagrangian models, it may be coupled to any gridded velocity fields, returning dispersal diagnostics as realistic as is the input flow field. Extensive description can be found in Rossi et al. (2014), Ser-Giacomi
et al. (2015) and Monroy et al. (2017). Here LFN simulates the dispersal of passively drifting propagules
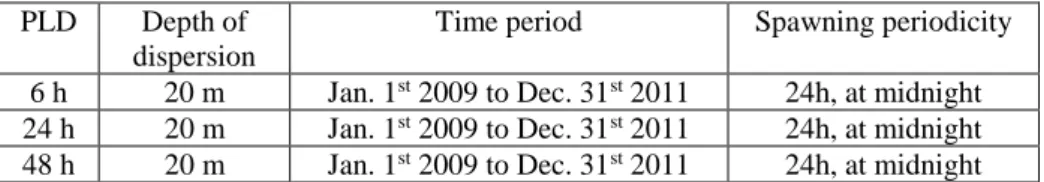
as horizontal Lagrangian trajectories obtained after integration of the high-resolution 2-dimensional flow field generated by the RHOMA configuration. Parameters are tuned in accord with both biological and numerical knowledge from the literature. Only the most relevant elements are summarized hereafter while the design of our numerical experiments are reported in Table 1.
To encompass all possible circulation schemes affecting dispersal, we simulate about 1090 starting dates representing successive spawning events occurring every day at midnight over years 2009-2011. Despite undetectable influence on our results (not shown), we retained this release time instead of midday as numerous benthic organisms spawn at dusk to limit mortality of their propagules by predation (Rasmuson et al. 2014). While the daily periodicity of spawning is higher than the prescriptions of Monroy et al. (2017), it provides large statistics to compute annual and seasonal means (see below). The tracking times mimic three different PLDs: 6, 24 and 48 hours. These PLDs would correspond to the dispersal traits of low-dispersing species inhabiting coralligenous grounds (see above). Despite the fact that some benthic species have planktotrophic larvae, supposedly surviving several days to a few weeks in the water column, the short time-scales of our study could still be relevant for these organisms as Cahill et al. (2017) reported genetic discontinuities in the bay for several species, including those dispersing via planktotrophic larvae. Note that PLDs longer than 48 hours are technically out-of-reach with this flow field as the mean residence time of water masses within the bay are usually of the same order of magnitude (ranging 1 to 5 days). In other words, tracking dispersal for durations longer than 48 hours with RHOMA would return biased results due to the limited extent of the domain (preventing the consideration of the influences of more distant patches).
The resolution of each quasi-squared node, representing a patch of coralligenous, is about 600 m (0.005°). Each node has the same area and contains approximately the same initial number of particles (proportional to the land-ratio); as prescribed by Monroy et al. (2017), 100 particles are seeded per full-ocean node. The time step of the Runge-Kutta algorithm is 10 min, fulfilling the Courant-Friedrichs-Lewy condition. We retained the vertical layer at 20 m, as it compromises the depth at which larvae of the target species are more likely to be released while it gives a good spatial overlap between the LFN grid and the coralligenous map. More specifically, the coastal mask of RHOMA is not consistent with the delimitation of coralligenous habitats of CARTHAM; as such, selecting a deeper layer of the model would force us to exclude many of the near coastal patches.
- Connectivity metrics and statistical analyses
For each experiment, LFN builds a connectivity matrix that encodes the movements of particles resulting from dispersal among all habitat patches. The diagonal elements represent the number of particles which stayed or returned to the same location after dispersal; the non-diagonal elements represent the particles exchanged between each pair of nodes, after dispersal during a given PLD, considering the origin (destination) node as a row (column, respectively) of the matrix. Thanks to various computations, including normalization procedure to get probabilities ranging from 0 to 1, we compute three connectivity diagnostics for each coralligenous patch following the formulation of Dubois et al. (2016). More specifically, Local Retention (LR) is computed as the ratio of particles retained in their origin node to all particles released from that node. Self-Recruitment (SR) is the ratio of particles retained in their origin node to all incoming particles (those that were released from this node as well as from the surrounding ones). It measures the proportion of local recruits that originated from the source population. SR differs from LR as it is a function of the number of propagules arriving from elsewhere while LR depends on the number of those leaving. Finally, we evaluate the relative importance of propagules export versus import by calculating the Source-Sink (SS) metric. It is the ratio of the ingoing number of propagules to the sum of both ingoing and outgoing propagules. To read more about the interpretation of such connectivity metrics and their robustness, the readers are referred to Dubois et al. (2016) and Monroy et al. (2017). The LFN methodology provides robust estimations of dispersal also for open fluid domains, as we study here (Ser-Giacomi et al. 2017).
Following Rossi et al. (2014) and Ser-Giacomi et al. (2015), we finally identify clusters of coralligenous patches as several groups of tighly-connected nodes. So-called “coralligenous provinces” are obtained by applying a state-of-art community detection algorithm called Infomap (Rosvall & Bergstrom, 2008). The partition is based on random walkers moving through the network with transition probabilities encoded in the connectivity matrix, an equivalent of network adjacency matrix. By exploiting the properties of information compression when describing such probability flow, Infomap finds the
optimal network partition when minimizing the mean size of the code-word that describes inter- and intra-community transitions. This methodology is especially suited to partition dispersal networks analyzed here as (i) it takes into account both “direction” and “fluxes” of all links, (ii) it does not constrain a-priori the number of communities, and (iii) it detects communities of different sizes, relieving the usual “resolution limit” typical of other clustering algorithms (Fortunato & Barthélemy, 2007). All in all, Infomap decomposes the network of coralligenous patches into an optimal number of communities, defining “coralligenous provinces” that are well connected internally but weakly connected among them. It allows us to analyze statistics of retention and exchanges of particles among synthetic provinces (Fig. 5 and Tab. 2).
We compute multi-annual means of connectivity diagnostics (LR, Fig. 2; SR, Fig. 3; SS, Fig. 4) and coralligenous provinces (Fig. 5) by averaging about 1090 daily experiments over 2009-2011, encompassing a total of more than 40 million Lagrangian trajectories. Seasonal means of LR and SS are obtained by averaging about 280 daily experiments (aggregating all experiments occurring during a given season across 3 years), encompassing more than 10 million Lagrangian trajectories. Annual means could be seen as relevant for modes of asexual (e.g. colony/algal fragmentation, sponge budding) or sexual reproduction that would occur all year long. Seasonal means is more relevant when the release of sexual or asexual propagules occurs during a specific season. Note that good statistics allow considering other manners to aggregate experiments in the future, for instance for species that have well-defined spawning period or whose spawning is triggered by predictable physical clues.
Results:
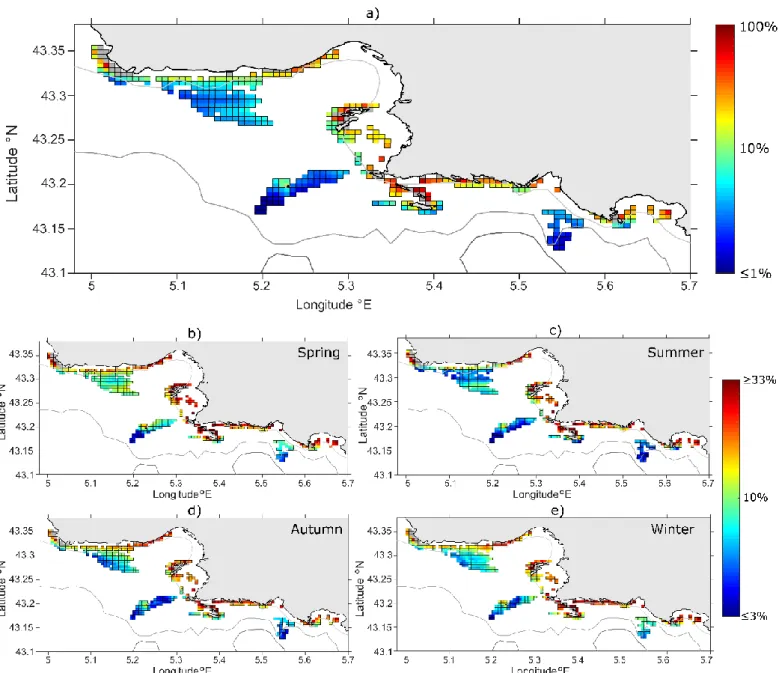
Values of Local Retention (LR) are highly dependent on the tracking time, with retention rates decreasing as the PLD increases (not shown). Total means (i.e. spatial average for 423 patches considering the multi-annual means) of LR are 10.2 % for PLD = 6 hours, 2.3 % for PLD = 24 hours and 1.5 % for PLD = 48 hours. When mapping annual mean of LR for a PLD of 6 hours (Fig. 2a), it reveals contrasting spatial patterns. Most coralligenous patches situated in the vicinity of the shoreline, i.e. within the 40 m isobaths, are usually characterized by retention rates ranging from 30 to 100 %. Conversely, the most offshore patches (e.g. around Planier island and off Cassis) are characterized by low retention, spanning 0-10 %. Note that retention around Planier islands is higher in its north-western side. LR exhibits substantial seasonal variability (Fig 2b to 2e) with highest total means simulated in spring (LR = 12.6 %) as compared to other seasons (ranging 8.8% to 9.6%). Minimum values are obtained for autumn and winter, suggesting more efficient and homogenous dispersion during these seasons. A few noticeable seasonal changes can be highlighted in Marseille bay: the retention along “Côte Bleue” is maximized in spring while showing local minima in summer; offshore patches (around Planier islands and off Cassis) are more retentive in winter and spring than in summer and autumn. Self-Recruitment (SR) also weakens when the PLD increases (Fig. 3) but to a lesser extent than LR: total means of SR are 17.5 % for PLD = 6 hours, 5.2 % for PLD = 24 hours and 3.3 % for PLD = 48 hours. Spatial patterns also reveal relatively high SR in shallower patches (ranging from 10 to 80%) as compared to most offshore locations where SR spans 0-10%. Conversely to LR, mean SR values along the Côte Bleue (from 5.05° to 5.2°E) and, to a lesser extent along the Calanques (from 5.4° to 5.5°E), tend to be weaker than along other coastlines for all PLDs. While LR values of all individual patches decrease when PLD increases, this does not hold for SR: shallower nodes tend to maintain moderate levels of SR even when the PLD increases (Fig. 3c).
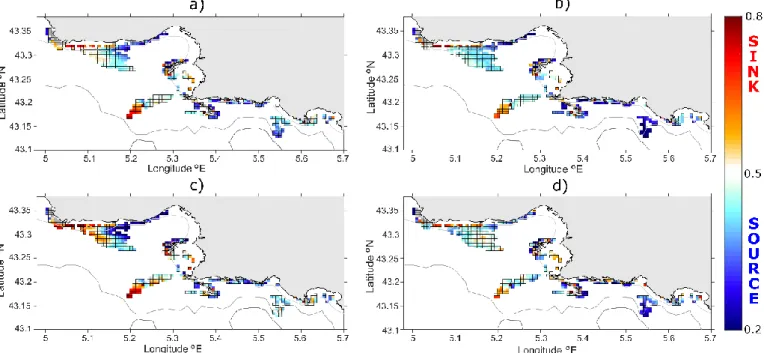
Source-Sink (SS) is less sensitive to the PLD than LR and SR (Fig. 4a, b). For all PLDs, patches off the western shorelines of Côte Bleue (from 5.05° to 5.18°E) are consistently characterized as sinks. Those located to the east and within the southern bay (from 5.18° to 5.3°E) are instead identified as sources. Most patches on the eastern side of the domain (Riou archipelago, Calanques, off Cassis and la Ciotat) are also mainly categorized as sources. Coralligenous patches around Planier islands show a bimodal pattern: sinks are identified on the offshore flank (e.g. south-westward) of the island while sources are
found on its inshore side (north-eastward). A few seasonal patterns can be distinguished (Fig. 4c, d): the marked sources and sinks previously described along Côte Bleue are emphasized during summer while the upstream sources (Riou archipelago, off Cassis) are weak in summer but maximized during winter.
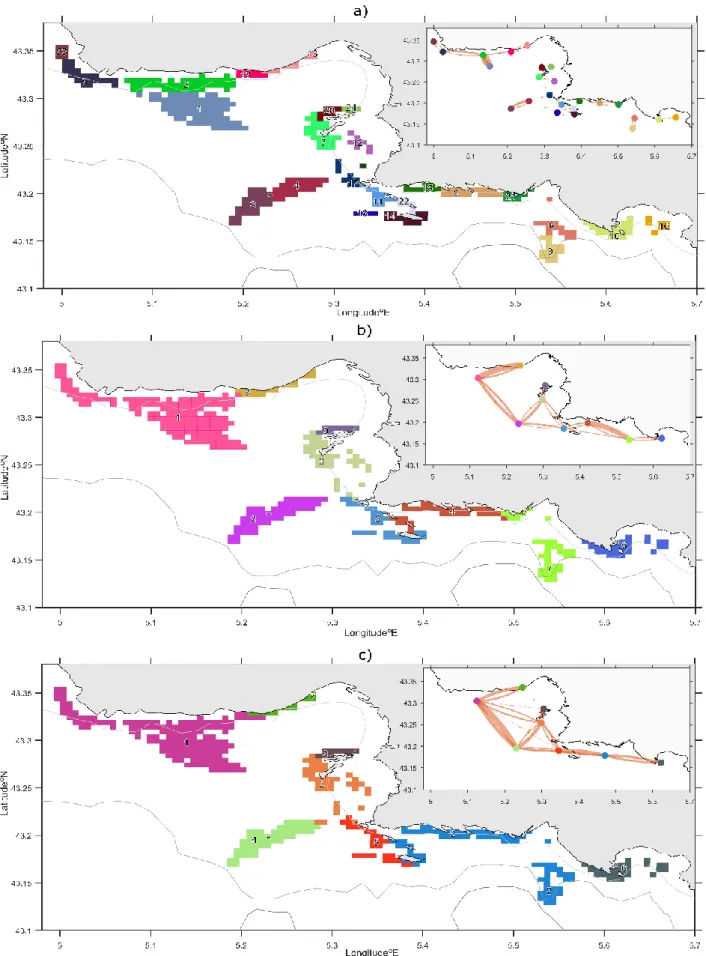
Clustered patches are highly dependent on the tracking duration: 24 coralligenous provinces are captured for a PLD of 6 hours while only 9 and 8 provinces were obtained for PLD of 24 and 48 hours, respectively (Fig. 5). The mean areas of these provinces scale with the PLD (Table 2): it is about 3.4 km2 for 6 hours, 9.1 km2 for 24 hours and 10.2 km2 for 48 hours. Regional local retention (i.e. similar to LR but computed at the “province” scale) is maximized by Infomap, reaching 90% or more for most provinces. Their exchanges with other provinces range several orders of magnitude (Tab. 2): for each province, a couple of links with neighboring provinces make up to 99% of the exchanges (plotted on the inserts of Fig. 5), while a few other links are weak and rare but do occur sometimes to time with more distant provinces (reported in Tab. 2). Focusing on the prominent links only, we find several subgroups of provinces which are disconnected for PLD of 6h. When the PLD increases, the provinces are less numerous and become larger; the network of coralligenous patches is more and more connected. Longer tracking times allow the development of bridging links among these provinces that were disconnected at short time-scales. For instance, the province around Planier Island is connected with the Côte Bleue province for PLD of 24 hours, while they were isolated at 6h. For 48h hours, the Planier/Côte Bleue connection remains while both Frioul and Riou archipelago act as intermediate hubs connecting relatively well the most distant patches located at both western (Côte Bleue) and eastern (Calanques, Cassis, La Ciotat) limits of the domain.
General discussion & conclusions:
- Relating mean patterns of habitat connectivity to hydrodynamics:
Our results suggest that retention processes are highly dependent on the bathymetry, e.g. elevated retention at sites where water depth is less than 20-40 m, due to weaker currents as compared to the more offshore locations (coastal currents tend to weaken by dissipative processes such as bottom friction). Intuitively, the longer dispersal time, the less retention of propagules. While this interpretation for LR is straightforward, it is not so valid for SR which combines together retained particles (numerator) and ingoing fluxes (denominator): so high values of SR can be caused by both high retention and/or low incoming fluxes, highlighting the counter-intuitive effect of recirculation processes in this semi-opened bay (which would not be prominent along straight coastlines). Indeed, LR and SR scale differently with the PLD probably because longer dispersal may allow some particles to leave a given patch and eventually return to it later on, emphasizing the importance of recirculation processes, especially in the southern bay.
Wind-induced circulation may be associated with the seasonal variations of LR off Côte Bleue: maxima occur in spring when SE/SW winds prevail, favoring coastal convergence explaining the coastal sinks, while minima are simulated during summer when NW winds and associated sporadic upwelling (i.e. coastal divergence, favoring coastal sources, as shown by Dubois et al. 2016) are prominent. The SS patterns described off Côte Bleue are emphasized during summer, probably due to the more frequent occurrence of wind-forced events during this season. More generally, sources are overall located at the eastern side of our domain and sinks at the western side (Fig. 4) due to the westward (cyclonic) global-scale circulation.
The relationship between the NC and our connectivity diagnostics is more puzzling. Earlier research suggested that the maximum transport of the NC, associated to a narrower jet flowing closer-to-shore, occur in winter (Alberola et al. 1995). The NC becomes baroclinically unstable especially in winter, producing large mesoscale meanders which may penetrate into the Gulf of Lion (Barrier et al. 2016) and Marseille Bay (Pairaud et al. 2011; Ross et al. 2016). This knowledge is consistent with the sinks
located to the south-east flank of Planier island and the weak sources associated with relatively high retention in the wake of the island (north-west side). The south-east sinks are likely receiving propagules from the identified upstream sources, e.g. Riou archipelago and patches off Cassis, which are weak in summer but maximized during winter. It is however contradictory with the seasonal minimum of LR simulated in summer/autum for the coralligenous habitats off Cassis, suggesting that NC intrusions may not impinge on these patches.
As the PLD increases, the provinces becomes larger. This is consistent with the monotonic increase of provinces areas with advection time already documented by Ser-Giacomi et al. (2015). The northern sector of Marseille Bay is weakly connected to Côte Bleue due to higher residence time (Pairaud et al. 2011). Currents at 20m depth are globally westwards along the Côte Bleue coastline (Fig. 4 and Fig. 5a). For PLD of 24h and 48h, Planier is connected with all the other provinces. Under the influence of all the above-mentioned major hydrodynamical processes (i.e. up/downwelling, NC meanders and intrusions and mesoscale eddies), Planier island acts as a major “connecting hub”. The transit times and pathways reported by Thibault et al. (2016), derived from the same RHOMA model using another methodology, are consistent with our analyses. For instance, they found that the connections between Planier and the Côte Bleue need a minimum of 12h to 18h to be realized, explaining why the corresponding provinces are disconnected for PLD = 6h while becoming connected for 24 and 48h. Note that our synthetic coralligenous provinces could also help identifying community similarities. Lagrangian particles could equally represent passive propagules or small water parcels; as such, pair of sites which connected at short time-scales (sufficiently short so that hydrographic conditions can be assumed to remain relatively constant) would also experience relatively similar abiotic conditions. Both processes could homogenize species diversity among local communities.
In fact, all hydrodynamical processes contribute altogether to the patterns of connectivity reported here; it is thus difficult to link a specific pattern with a given process. For instance, SR values along the Côte Bleue (from 5.05° to 5.2°E) and, to a lesser extent along the Calanques (from 5.4° to 5.5°E), tend to be weaker than other coastlines. It may be because local propagules spread offshore due to upwelling events while recirculation processes related to NC intrusions on one hand, and to the occurrence of the “Marseille Eddy” on the other hand, would bring particles from elsewhere into Calanques and Côte Bleue, respectively (Schaeffer et al. 2011; Fraysse et al. 2014). Moreover, as hydrodynamics exhibit high spatio-temporal variability, large statistics are required to get robust diagnostics of connectivity encompassing all processes. The good ability of RHOMA to reproduce realistically the complex circulation of Marseille Bay over 2009-2011 reported by Fraysse et al. (2013) suggest that our average patterns are robust while taking into account variability. Note however that inter-annual variability is important in the NW Mediterranean (Hidalgo et al. 2019). Indeed year 2009 was characterized by unusual conditions: a long-lasting summer warming event occurs down to 40 m, inducing mortality for
P. Clavata populations (Pairaud et al. 2014) and possibly exceptional circulation patterns. Nevertheless,
the exact dispersal pathways of water parcels originating from the Calanques studied by Millet et al. (2018) for two very distinct NC intrusion events (in june 2008 and October 2011) are consistent with our mean patterns. As the circulation of the bay depends on short wind-forced events (dominated by north-west and south-east winds, Millet et al. 2018) occurring episodically at all time of the year, future work could aim at producing climatology of “wind-forced events”. By categorizing all events characterized by similar wind-forcing conditions across the 3-year period and by aggregating matrices accordingly (independently of the season and the year), one could describe extreme connectivity. It would contrast the mean patterns analyzed here and could be relevant for species whose spawning is triggered by specific physical clues such as wind-forced upwelling and associated coastal cooling. Note however that wind-driven currents are often associated with strong vertical velocities that are neglected in our “horizontal” assumption. Other perspectives could be to analyze other dispersal depth and to perform 3-dimensional simulations.
- Discussing the biological implications of our simulated results:
According to Dubois et al. 2016, the integrated analyses of our metrics inform on the connectivity of coralligenous provinces. When both LR and SR are high, as in both northern and southern sectors of Marseille Bay, it symbolizes mostly “closed” populations for which one could expect high differentiation with external sites and low diversity. The Côte Bleue also displays consistent patterns of retention: both LR and SR are large along both eastern and western domains (from 5° to 5.05°E and from 5.2° to 5.3°E) and moderate in-between (5.05-5.2°E). SS shows a discontinuity at about 5.2°E, with sinks westward and sources eastward. Altogether, spatial patterns of LR, SR and SS suggest (i) low diversity along Côte Bleue and (ii) a sharp change of connectivity at about 5.2°E, which was also the presumed north-west limit of the Calanques influence (Millet et al. 2018).
Interestingly, a multi-specific population genetics study along French Mediterranean shorelines (especially detailed in Marseille Bay) confirms the prediction of a low genetic diversity for Côte Bleue: for most species, it has a significant negative contribution to global genetic diversity (Cahill et al. 2017).
While they invoked anthropogenic activities, such as habitat degradation or pollution, to explain variations in local genetic diversity, the present study suggests that physical connectivity among patches could alone explain their observations. For the builder algae L. stictiforme, the Côte Bleue population (Carry-le-Rouet and Couronne) actually exhibits the lowest genetic diversity as compared to the populations sampled around Frioul and Riou archipelago as well as Cassis (De Jode, 2018). Monospecific population genetic studies comparing sites across the ~5.2°E discontinuity tend to support its effect as a barrier to gene flow. For Cystoseira amentacea, an algae of the shallowest infra-littoral not restricted to coralligenous habitats, Thibaut et al. (2016) reported genetic structuring concordant with our results: among the Cote Bleue samples, Niolon population (5.25°E) appear well differentiated from the most western sampling sites (Carro and Ponteau). For the red coral C. rubrum, Ledoux et al. (2010) found significant differentiation between the population situated at about 5.2°E and the other populations sampled at Carro (5.04°E) and Ponteau (~5°E).
Conversely, when both LR and SR are low, as for Planier Island and Cassis provinces, it suggests largely “open” population with high genetic diversity. Furthermore, the offshore flank of Planier province is a sink (i.e. net importer of external propagules with little retention of its own propagules), suggesting it could house higher diversity than Cassis, which behaves as a source (i.e. net exporter). While these conclusions seem robust for Planier Island, the behavior of Cassis could change if the model domain was larger, hence allowing us to consider larval influences from upstream patches (i.e. further east, from “côte Varoise”). The high diversity expected at Planier Island, also supported by its central position in the network (Fig. 5), is consistent with Thibault et al. (2016) who found that Planier Island is a mix of all origins with high genetic diversity. This central hub ensure the links between the most western and eastern patches at 24h and 48h; it could also ensure such connection at 6h thanks to stepping-stone processes.
The networks of coralligenous provinces obtained at different time-scales provide insights about the kind of biological clusters one should expect if habitat connectivity is the main factor structuring population. Thibaut et al. (2016) found a strong and significant genetic structure within Marseille Bay for C. amentacea, whose dispersal is ensured by a zygote with survival time spanning 4-18h. Pairs of sites characterized by low genetic differentiation are those tightly connected by ocean currents, as indicated by the synthetic provinces obtained for 6h (Fig. 5a). Cahill et al. (2017) also found that the most important barriers of genetic differentiation separate Côte Bleue from the rest of the Bay. The crustose coralline algae of the genus Lithophyllum (De Jode et al. 2019) or the bryozoan M. truncata (De Jode, 2018), which are both engineering species of coralligenous, exhibited genetic differentiation in accord with our results. These species have propagules dispersing from a few hours to a day, explaining why their genetic structures match well our model results for 6h and 24h (Fig. 5a, b). Observed structures of the red coral documented significant genetic differentiations between Côte Bleue, Marseille Bay and Calanques (Ledoux et al. 2010, Pratlong et al. 2018), suggesting effective
dispersal closer to 6h than to 24h. Note also that our provinces and their typical spatial-scales could be used to guide future sampling efforts (Dubois et al. 2016), e.g. by targeting disconnected provinces while avoiding duplicate sampling in each province.
An important aspect of the present approach is that it constrains current-driven dispersal by considering habitat patchiness in order to estimate biological connectivity. Although population genetics stipulate that gene flow is proportional to the product of ‘migration rate’ and ‘effective population size’ (genetic drift), most studies interpreting genetic structures invoke the duration of dispersal solely, without discussing the effective population size. The latter should be, at first order, proportional to the overall extent of the preferential habitat of the species. Although gorgonians have similar dispersal traits, genetic differentiation in the bay was lower in E. cavolini than in P. clavata, and lower in P. clavata than in the red coral C. rubrum (Canovas-Molina et al., 2018). The respective spatial extension of their actual habitats may explain the different levels of genetic differentiation: E. cavolini is widespread over various habitats (not restricted to coralligenous) while C. rubrum displays the less extended populations, exclusively developing on coralligenous patches. So even if the species of interest is restricted, or not, to coralligenous habitats, our synthetic provinces incorporating both habitat patchiness and current-driven connectivity may bring interesting insights into population structure.
Species community differentiation (e.g. beta-diversity) could be affected by connectivity in a similar way as genetics: well-connected communities would show more similarities. In Marseille bay, species diversity varies from site-to-site but, contrary to intra-specific studies, there is no clear spatial structuring within the bay (De Jode, 2018; Doxa et al. 2016; Cinar et al. 2020). Local abiotic factors seem to control community composition at small scales more likely through natural selection (De Jode, 2018), reflecting favourable ecological niches rather than dispersal limitations (conversely to “within-species” structuring). Nevertheless, the present study could help explaining community composition under certain circumstances, such as when mortality events affect a population. In such cases, their transient spatial distributions would follow recolonization routes, which result from both ocean currents and favourable abiotic conditions to settle. After the P. clavata mortality outbreak in summer 2009, Mokhtar-Jamai et al. (2011) observed significant genetic structure in Marseille bay, including a genetic break around Cap Croisette. It matches best with our coralligenous provinces defined for 6 hours, despite its lecithotrophic larvae potentially dispersing longer. Investigating population structures of genus Echinocardium (sand-dweller sea urchins that have planktotrophic larva dispersing for about 3 weeks), Egea (2011) observed that E. cordatum is prominent within Marseille bay till the western flanks of Riou archipelago (~5.4°E); it is then replaced by E. mediterraneum in “Calanques” and along eastern coastlines. Only the 6-hour synthetic provinces reveal a connectivity break at 5.4°E, while it vanishes for longer dispersal. It might indicate that the biogeography of Echinocardium is primarily controlled by abiotic factors instead of dispersal. Overall, it suggests that short connectivity (< 12h) in this region tend to be associated with relatively constant abiotic conditions whereas longer connectivity (≥ 24-48h) would overcome stronger environmental gradients, potentially preventing propagules to survive along dispersal routes and/or to settle at final destination.
- General conclusions:
We investigated fine-scale connectivity of coralligenous habitats in Marseille bay for a range of parameters relevant for many benthic species using dispersal modelling and high-resolution cartography. We analyzed annual and seasonal averages of three connectivity metrics measuring the retention and exchange of propagules driven by ocean currents within and among habitat patches. Our synthetic coralligenous provinces and associated statistics for several time-scales provide broader views of the ecological clusters structuring benthic populations. By discussing our results against genetic and ecological studies, we emphasized previous findings that are well supported by our simulated habitat connectivity. Altogether, it suggests that current-driven dispersal of propagules and habitat patchiness control to a certain degree the spatial structure of benthic populations.
It illustrates the potential of such interdisciplinary studies to reveal counter-intuitive and non-straightforward relationships among hydrodynamics and population structures that could worth further investigations. Moreover, our simulated connectivity patterns could be used by other researchers to design future sampling and to help explaining observed population structures. Last but not least, model outputs are available upon request and can be aggregated at other levels, such as monthly/weekly time-scales or as a climatology of wind-forced events.
Among the conclusions backed up by both modelling and observations, our study suggests that low diversity is expected along Côte Bleue whereas Planier Island, which acts as a key connecting hub for all surrounding coralligenous patches, would be characterized by high diversity. In combination with an index evaluating the conservation state of coralligenous habitats (e.g. Sartoretto et al. 2017), these areas could be considered as good candidates for constant monitoring and protection (Ods et al. 2012). Future work may focus on how habitat connectivity will evolve with climate change (Gerber et al. 2014) and with the constant development of anthropogenic structures such as harbors, wind farms and offshore platforms (Henry et al. 2018).
Acknowledgements:
V.R, M.L., T.L. and A.C acknowledge financial support from the European project SEAMoBB, funded by ERA-Net Mar-TERA and managed by ANR (number ANR_17_MART-0001_01, P.I.: A.C.). T.L. is funded by a MRT doctoral fellowship obtained through Aix-Marseille University. The authors acknowledge technical assistance and access to high-performance computing facilities provided by the IT staff of the ‘OSU Institut Pytheas’ (Aix-Marseille University, INSU-CNRS). J.G. Harmelin kindly provided his bibliographic advice. M. Verlaque and T. Thibaut are thanked for their assistance with algal taxonomy. E. Renard and A. Ereskovsky informed us on Sponge reproductive strategies. We are also grateful to “Andromède Oceanology” and “Agence de l'Eau” for providing us the high-resolution coralligenous cartography from the online platform Medtrix.
References:
Alberola C, Millot Claude, Font J (1995). On the seasonal and mesoscale variabilities of the Northern Current during the PRIMO-0 experiment in the western Mediterranean-sea. Oceanologica Acta, 18(2), 163-192.
Andromede Oceanologie (2014). La Méditerranée dévoile ses dessous - Cartographie continue des habitats marins. Partenariat Agence de l'eau RMC - Andromède.
Astruch P., Goujard A., Charbonnel E., Rogeau E., Rouanet E., Bachet F., Bricout R., Bonhomme D., Antonioli P. A., Bretton O., Monin M., Harmelin J.G., Sartoretto S., Chevaldonne P., Zibrowius H., Verlaque M., 2011. Inventaires biologiques et Analyse écologique de l’existant, Natura 2000 en mer, Lot n°12 « Côte Bleue Marine » FR 9301999 ; GIS Posidonie & Agence des Aires Marines Protégées, GIS Posidonie/PMCB publ., 400 p.
Astruch P., Rouanet E., Chemisky B., Seguin E., Bonhomme D., Goujard A., Le Direach L., Harmelin J.G., Vacelet J., Perez T., Chevaldonne P., Bellan G. (2012). Inventaires biologiques et analyse écologique de l’existant, Natura 2000 en mer, Lot n°6 « Calanques et îles marseillaises – Cap Canaille et Massif du Grand Caunet » FR 9301602. Contrat COMEX SA/GIS Posidonie – Agence des Aires Marines Protégées, COMEX SA/GIS Posidonie publ., 500 p.
Ballesteros E. (2006). « Mediterranean coralligenous assemblages: a synthesis of present know- ledge ». In : Oceanography and Marine Biology : An Annual Review 44, p. 123–195.
Barrier, N., AA Petrenko, Y Ourmieres (2016) Strong intrusions of the Northern Mediterranean Current on the eastern Gulf of Lion: insights from in-situ observations and high resolution numerical modelling, Ocean Dynamics, 66 (3), 313-327.
Botsford, L.W., Brumbaugh, D.R., Grimes, C. et al. (2009) Connectivity, sustainability, and yield: bridging the gap between conventional fisheries management and marine protected areas. Rev Fish Biol Fisheries 19, 69–95.
Cahill AE, De Jode A, Dubois S, Bouzaza Z, Aurelle A, Boissin E, Chabrol O, David R, Egea E, Ledoux J-B, Mérigot B, Weber A A-T, and Chenuil A. (2017). A multispecies approach reveals hot-spots and cold-spots of diversity and connectivity in species with contrasting dispersal modes. Molecular Ecology, 26, 6563-6577.
Cánovas-Molina A, Montefalcone M, Bavestrello G, Masmoudi MB, Haguenauer A, Hammami P, Chaoui L, Kara MH and Aurelle D (2018). From Depth to Regional Spatial Genetic Differentiation of Eunicella Cavolini in the NW Mediterranean. Comptes Rendus Biologies 341: 421-32.
Cinar M.E., Féral J.-P. et al. (2020). Coralligenous assemblages along their geographical distribution: testing of concepts and implications for management, Aquatic Conservation: Marine and Freshwater Ecosystems, in press.
Costantini, F., Ferrario, F., & Abbiati, M. (2018). Chasing genetic structure in coralligenous reef invertebrates: patterns, criticalities and conservation issues. Scientific Reports, 8(1), 5844. doi: 10.1038/s41598-018-24247-9
Crisci, C., Bensoussan, N., Romano, J. C., & Garrabou, J. (2011). Temperature anomalies and mortality events in marine communities: insights on factors behind differential mortality impacts in the NW Mediterranean. PLoS One, 6(9).
De Jode, A., David, R., Haguenauer, A., Cahill, A. E., Erga, Z., Guillemain, D., … Chenuil, A. (2019). From seascape ecology to population genomics and back. Spatial and ecological differentiation among cryptic species of the red algae Lithophyllum stictiforme/L. cabiochiae, main bioconstructors of coralligenous habitats. Molecular Phylogenetics and Evolution, 137, 104–113.
De Jode, A. (2018). Etude de la biodiversité des habitats coralligènes et de l'influence des facteurs environnementaux par des approches génétiques: des populations d'espèces ingénieures aux communautés (Doctoral dissertation, Aix-Marseille).
Doxa, A., Holon, F., Deter, J., Villéger, S., Boissery, P., & Mouquet, N. (2016). Mapping biodiversity in three-dimensions challenges marine conservation strategies: The example of coralligenous assemblages in North-Western Mediterranean Sea. Ecological Indicators, 61, 1042–1054. doi: 10.1016/j.ecolind.2015.10.062
Dubois, M., V. Rossi, E. Ser-Giacomi, S. Arnaud-Haond, C. López and E. Hernández-García, (2016). Linking basin-scale connectivity, oceanography and population dynamics for the conservation and management of marine ecosystems, Global Ecology and Biogeography, 25, 503–515.
Duputie, A., and Massol, F. (2013). An empiricist’s guide to theoretical predictions on the evolution of dispersal. Interface Focus, 3: 20130028.
Egea, E. (2011). Histoire évolutive, structures génétique, morphologique et écologique comparées dans un complexe d’espèces jumelles : Echinocardium cordatum (Echinoidea, Irregularia). http://www.theses.fr. Thesis, Aix-Marseille 2. 321pp.
Ferretti, C., Magnino, G., & Balduzzi, A. (2007). Morphology of the larva and ancestrula of Myriapora truncata (Bryozoa, Cheilostomatida). Italian Journal of Zoology, 74(4), 341–350.
Fortunato S. and M. Barthelemy (2007). Resolution limit in community detection, Proc. Natl. Acad. Sci. U.S.A. 104, 36–41.
Fraysse M., C. Pinazo, V. Faure, R. Fuchs, P. Lazzari, P. Raimbault, and I.L. Pairaud, (2013). 3D coupled physical-biogeochemical model development around Marseille’s coastal area (NW Mediterranean Sea): What complexity is required in coastal zone? Plos One, 8 (12): e80012. doi:10.1371/journal.pone.0080012.
Fraysse, M., I. Pairaud, O. N. Ross, V. M. Faure, and C. Pinazo (2014). Intrusion of Rhone River diluted water into the Bay of Marseille: Generation processes and impacts on ecosystem functioning, J. Geophys. Res. Oceans, 119, 6535–6556.
Gerber, L. R., M. Del Mar Mancha-Cisneros, M. I. O’Connor, and E. R. Selig. (2014). Climate change impacts on connectivity in the ocean: Implications for conservation. Ecosphere 5(3):33. http://dx.doi.org/10.1890/ES13-00336.1
Garrabou, J. and E. Ballesteros (2000). « Growth of mesophyllum alternans and lithophyllum frondosum (corallinales, rhodophyta) in the northwestern mediterranean ». In : European Journal of Phycology 35.1, p. 1–10.
Garrabou, J., Coma, R., Bensoussan, N., Bally, M., Chevaldonné, P., [...], Ledoux, J. B. (2009). Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global change biology, 15(5), 1090-1103.
Goffredo, S., Marchini, C., Rocchi, M., Airi, V., Caroselli, E., … Zaccanti, F. (2012). Unusual pattern of embryogenesis of Caryophyllia inornata (scleractinia, caryophylliidae) in the mediterranean sea: Maybe agamic reproduction? Journal of Morphology, 273(9), 943–956. doi: 10.1002/jmor.20039.
Henry, L., Mayorga-Adame, C.G., Fox, A.D. et al. (2018). Ocean sprawl facilitates dispersal and connectivity of protected species. Sci Rep 8, 11346. https://doi.org/10.1038/s41598-018-29575-4
Hidalgo M., V. Rossi et al., 2019. Accounting for ocean connectivity and hydroclimate in fish recruitment fluctuations within transboundary metapopulations. Ecological Applications, 29(5), e01913, doi:10.1002/eap.1913.
Jones, G.P., M. Srinivasan and G.R. Almany (2007) Population Connectivity and Conservation of Marine Biodiversity, Oceanography, vol. 20, No. 3, pp. 100-111.
Lazure P. and Dumas F. (2008). An external-internal mode coupling for a 3D hydrodynamical model for applications at regional scale (MARS). Advances In Water Resources, 31(2), 233-250.
Ledoux, J.-B., Mokhtar‐Jamaï, K., Roby, C., Féral, J.-P., Garrabou, J., & Aurelle, D. (2010). Genetic survey of shallow populations of the Mediterranean red coral [Corallium rubrum (Linnaeus, 1758)]: new insights into evolutionary processes shaping nuclear diversity and implications for conservation. Molecular Ecology, 19(4), 675–690. doi: 10.1111/j.1365-294X.2009.04516.x
Martin, C., Giannoulaki, M., De Leo, F. et al. (2015). Coralligenous and maërl habitats: predictive modelling to identify their spatial distributions across the Mediterranean Sea. Sci Rep 4, 5073, https://doi.org/10.1038/srep05073.
Martin, S., A. Charnoz, and J.-P. Gattuso (2013a). Photosynthesis, Respiration and Calcification in the Mediterranean Crustose Coralline Alga Lithophyllum Cabiochae (Corallinales, Rhodophyta). European Journal of Phycology, 48(2), 163-72.
Martin, S., S. Cohu, C. Vignot, G. Zimmerman et J.-P. Gattuso (2013b). One-year experiment on the physiological response of the Mediterranean crustose coralline alga, Lithophyllum cabiochae, to elevated pCO2 and temperature. Ecology and Evolution, 3(3), p. 676– 693.
Martínez-Quintana, A., Bramanti, L., Viladrich, N., Rossi, S., & Guizien, K. (2015). Quantification of larval traits driving connectivity: the case of Corallium rubrum (L. 1758). Marine Biology, 162(2), 309–318. doi: 10.1007/s00227-014-2599-z
Millet B., Pinazo C., Banaru D., Pagès R., Guiart P., Pairaud I. (2018) Unexpected spatial impact of treatment plant discharges induced by episodic hydrodynamic events: Modelling Lagrangian transport of fine particles by Northern Current intrusions in the bays of Marseille (France). PLoS ONE 13(4): e0195257.
Mokhtar-Jamaï, K., Pascual, M., Ledoux, J.-B., Coma, R., Féral, J.-P., Garrabou, J., & Aurelle, D. (2011). From global to local genetic structuring in the red gorgonian Paramuricea clavata: the interplay between oceanographic conditions and limited larval dispersal.
Molecular Ecology, 20(16), 3291–3305.
Monroy, P., V. Rossi, E. Ser-Giacomi, C. Lopez, E. Hernandez-Garcia, (2016). Sensitivity and robustness of larval connectivity diagnostics obtained from Lagrangian Flow Networks, ICES Journal of Marine Science, doi:10.1093/icesjms/fsw235.
Nicolle A., Garreau P, Liorzou B (2009). Modelling for anchovy recruitment studies in the Gulf of Lions (Western Mediterranean Sea).Ocean Dynamics, 59, 953–968.
Norton, T. A. (1992). Dispersal by macroalgae. British Phycological Journal, 27(3), 293–301. doi: 10.1080/00071619200650271
Olds, A.D., Connolly, R.M., Pitt, K.A. and Maxwell, P.S. (2012), Habitat connectivity improves reserve performance. Conservation Letters, 5: 56-63. doi:10.1111/j.1755-263X.2011.00204.x
Opazo, L. F., & Otaíza, R. D. (2007). Vertical distribution of spores of blade-forming Sarcothalia crispata (Gigartinaceae) and crustose corallines (Corallinaceae) in the water column. Botanica Marina, 50(2). doi: 10.1515/BOT.2007.011
Pairaud, I., J. Gatti, N. Bensoussan, R. Verney and P. Garreau (2011). Hydrology and circulation in a coastal area off Marseille: Validation of a nested 3D model with observations. Journal of Marine Systems 88 (2011) 20–33.
Pairaud,I., Bensoussan,N., Garreau,P., Faure,V., Garrabou,J., (2014). Impacts of climate change on coastal benthic ecosystems: assessing the current risk of mortality outbreaks associated with thermal stress in NW Mediterranean coastal areas. Ocean Dyn. 64 (1), 103-115. Piazzi, L., Gennaro, P., & Balata, D. (2012). Threats to macroalgal coralligenous assemblages in the Mediterranean Sea. Marine pollution bulletin, 64(12), 2623-2629.
Pratlong, M., Haguenauer, A., Brener, K., Mitta, G., Toulza, E., Garrabou, J., Bensoussan, N., Pontarotti P., & Aurelle, D. (2018). Separate the wheat from the chaff: genomic scan for local adaptation in the red coral Corallium rubrum. bioRxiv, 306456, ver. 3 peer-reviewed and recommended by PCI Evol Biol.
Rasmuson, LK; Couture, JL; Morgan, SG; 2014. Weakly synchronized larval release maintained in the presence of predatory fishes. Journal of experimental biology and ecology, 454: 26-31.
Rodriguez-Prieto, C. (2016). Light and temperature requirements for survival, growth and reproduction of the crustose coralline Lithophyllum stictaeforme from the Mediterranean Sea, Botanica Marina, 59, 95-104.
Ross, O.N., M. Fraysse, C. Pinazo, I. Pairaud, (2016). Impact of an intrusion by the Northern Current on the biogeochemistry in the eastern Gulf of Lion, NW Mediterranean. Estuarine and Continental Shelf Research,170, 1-9, doi: 10.1016/j.ecss.2015.12.022
Rossi, V., E. Ser-Giacomi, M. Dubois, P. Monroy, M. Hidalgo, E. Hernandez-Garcia, C. Lopez, (2016). Lagrangian Flow Networks: a new framework to study the multi-scale connectivity and the structural complexity of marine populations, Proceedings (Ed. F. Briand) of the CIESM International Research Workshop n.48 entitled "Marine Connectivity - Migratory routes, stepping stones, larval dispersal", Soller (Mallorca), Spain.
Rossi, V., E. Ser-Giacomi, C. Lopez and E. Hernandez-Garcia, (2014). Hydrodynamic provinces and oceanic connectivity from a transport network help designing marine reserves, Geophysical Research Letters, 41 (8), 2883–2891.
Rosvall, M., and C. T. Bergstrom (2008), Maps of random walks on complex networks reveal community structure, Proc. Natl. Acad. Sci. U.S.A., 105(4), 1118–1123.
Schaeffer, A., Molcard, A., Forget, Ph., Fraunié, Ph., Garreau, P., (2011). Generation mechanisms for mesoscale eddies in the Gulf of Lions: radar observation and modeling, Ocean Dynamics, 61:1587–1609.
Sartoretto, S., T. Schohn, C.N. Bianchi, C. Morri, J. Garrabou, E. Ballesteros, S. Ruitton, et al. (2017). An integrated method to evaluate and monitor the conservation state of coralligenous habitats: The INDEX-COR approach, Marine Pollution Bulletin, 120, 222-231. Ser-Giacomi, E., V. Rossi, C. López and E. Hernández-García, (2015). Flow networks: A characterization of geophysical fluid transport, Chaos, 25 (3).
Ser-Giacomi, E., V Rodriguez-Mendez, C Lopez, E Hernandez-Garcia (2017). Lagrangian Flow Network approach to an open flow model, The European Physical Journal Special Topics, 226 (9), 2057–2068.
Thibaut T., L. Bottin, D. Aurelle, C.-F. Boudouresque, A. Blanfuné, M. Verlaque, I. Pairaud and B. Millet (2016). Connectivity of populations of the seaweed Cystoseira amentacea within the Bay of Marseille (Mediterranean Sea): genetic structure and hydrodynamic connections. Cryptogamie, Algologie, 2016, 37 (4): 233-255.
Thierry de Ville d'Avray, L. (2018). Identification and valuation of ecosystem services provided by coralligenous habitats. Doctoral dissertation, Aix Marseille Université, 261 p.
Thierry de Ville d’Avray L., Ami, D., Chenuil, A., David, R., & Féral, J. P. (2019). Application of the ecosystem service concept at a small-scale: The cases of coralligenous habitats in the North-western Mediterranean Sea. Marine pollution bulletin, 138, 160-170. Torrents, O., J. Garrabou, C. Marschal and J. G. Harmelin (2005). Age and size at first reproduction in the commercially exploited red coral Corallium rubrum (L.) in the Marseilles area (France, NW Mediterranean), Biological Conservation, 121, 391-97.
Figure 1: maps of the study area. Panel (a) represents the domain covered by the RHOMA hydrodynamical model
(black dots symbolize the coastal mask at 20 m); annotations indicate the names of a few specific locations recalled in the main text; the red dotted rectangle highlights the core region of study (including, from west to east, “Côte
Bleue”, “bay of Marseille”, “Calanques” and “La Ciotat”). Panel (b) displays the CARTHAM map of coralligenous habitats (dark red dots) with the LFN grid superimposed (semi-transparent rectangles colored according to their land-ratio, reddish colors indicate full ocean nodes while yellowish colors stand for nodes partially covered by land).
Figure 2: maps of Local Retention (LR, in % using log-scale) for a PLD of 6 hours. Panel (a) represents the multi-annual
mean (over 2009-2011); lower panels are seasonal averages: (b) spring, (c) summer, (d) autumn, (e) winter. The grey-colored nodes have been disregarded as they are partially- or fully-covered by land, resulting in a poor reliability of LR values in these nodes. The grey-scaled lines represent the 40 m (clearest), 100 m and 200 m (darkest
Figure 3: maps of Self-Recruitment (SR, in % using log-scale). Multi-annual mean (over 2009-2011) are displayed for
various PLDs: panel (a) is 6 hours, panel (b) is 24 hours and panel (c) is 48 hours. The grey-colored nodes have been disregarded as they are partially- or fully-covered by land, resulting in a poor reliability of SR values in these nodes.
Figure 4: maps of Source-Sink (SS, near-zero values indicate “sources”; values approaching 1 represent “sinks”).
Multi-annual mean (over 2009-2011) are displayed for PLD = 24 hours in panel (a) and for PLD = 48 hours in panel (b). Seasonal averages for PLD = 24 hours are displayed for summer in panel (c) and for winter in panel (d). The
grey-colored nodes have been disregarded as they are partially- or fully-covered by land, resulting in a poor reliability of SS values in these nodes. The grey-scaled lines represent the 40 m (clearest), 100 m and 200 m (darkest line)
Figure 5: maps of clustered coralligenous patches. Synthetic provinces are derived from the multi-annual mean
connectivity matrices for PLD = 6h in panel (a), for PLD =24h in panel (b) and for PLD=48h in panel (c). In all panels, each province is identified with a specific color (randomly chosen) and a number (consistent with those reported in
table 2). Upper right inserts symbolize the averaged network of provinces (colored dots) and their main directed links (reddish arrows; width is proportional to the fluxes of particles). It excludes both the exchanges lower than 1% and the regional local retention (reported in Tab. 2). The grey-scaled lines represent the 40 m (clearest), 100 m and
PLD Depth of dispersion
Time period Spawning periodicity
6 h 20 m Jan. 1st 2009 to Dec. 31st 2011 24h, at midnight
24 h 20 m Jan. 1st 2009 to Dec. 31st 2011 24h, at midnight
48 h 20 m Jan. 1st 2009 to Dec. 31st 2011 24h, at midnight
PLD (hours) Province identity Area (km2) Regional Local Retention (%)
Identity of connected provinces
< 0.1 % 0.1 ≤ x < 1 % ≥ 1% (inserts Fig. 6) 6 1 15.6 95 5 - 2 2 9 88.8 - 19 1 ; 5 3 5.8 95.4 - - 4 4 5.4 87.8 1 ; 18 7 ; 17 3 5 4.4 94 - 1 2 ; 23 6 2.8 94.6 8 10 ; 21 9 7 4.8 94.8 4 ; 24 17 12 ; 20 8 3.6 92.7 - - 15 ; 21 9 3 89.2 21 - 6 10 4.2 98.5 - 6 ; 16 - 11 2.2 95.6 - - 14 ; 17 ; 18 12 1.6 98.8 - 7 ; 17 - 13 2.8 93.3 2 - 19 14 2.8 89.1 4 ; 8 ; 15 ; 17 22 11 ; 18 15 1.8 97.8 - 8 22 16 1.4 96.4 - - 10 17 2 94.3 12 11 ; 18 4 ; 7 18 0.8 82.1 - 3 ; 17 4 ; 11 ; 14 19 1.2 76.8 - - 2 ; 13 ; 19 20 1.4 86.7 - - 7 ; 24 21 1.6 74.5 - 6 8 22 1.4 96.2 - 15 14 23 1.2 83.2 - - 5 24 0.8 95.5 - - 20 24 1 30.4 96.7 5 ; 9 3 2 ; 8 2 11.2 76.9 8 9 1 ; 3 ; 5 3 7.8 90.6 4 ; 8 1 2 ; 5 ; 9 4 7.2 92.5 2 ; 6 - 5 ; 7 5 6.4 79.9 1 ; 9 7 2 ; 3 ; 4 6 5.6 93.6 - 4 ; 5 7 7 7.2 56.7 - 2 4 ; 5 ; 6 8 4.4 64.7 2 ; 3 ; 9 - 1 9 2 74.5 8 1 ; 2 3 48 1 31.6 92.8 2 ; 5 8 3 ; 4 ; 7 2 15.8 84.6 7 ; 8 1 ; 3 4 ; 5 ; 6 3 8 78.4 - 2 ; 7 1 ; 4 ; 5 ; 8 4 10.8 41.5 - 2 ; 7 1 ; 3 ; 4 ; 5 ; 8 5 5.2 65.6 6 ; 7 8 1 ; 2 ; 3 ; 4 6 5.6 82.1 1 ; 3 4 ; 5 2 7 3.2 59.9 4 3 ; 8 1 8 2 68.4 5 4 1 ; 3 ; 7
Table 2: Statistics of exchange and retention among the synthetic coralligenous provinces