Colour and fluorescence emission of Euchroea auripigmenta beetle
Texte intégral
Figure

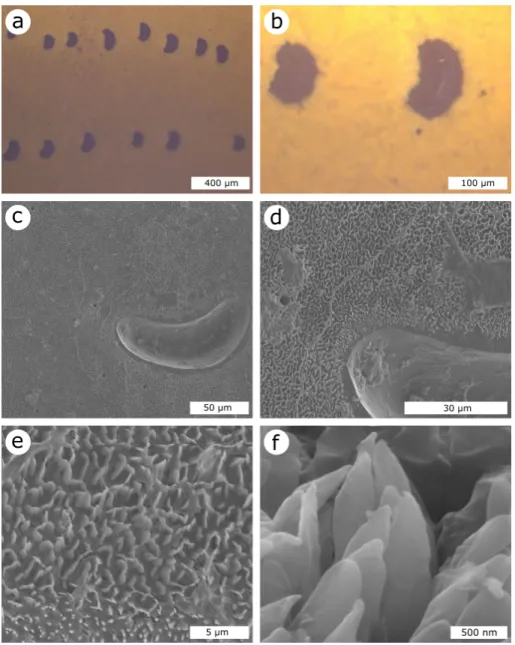

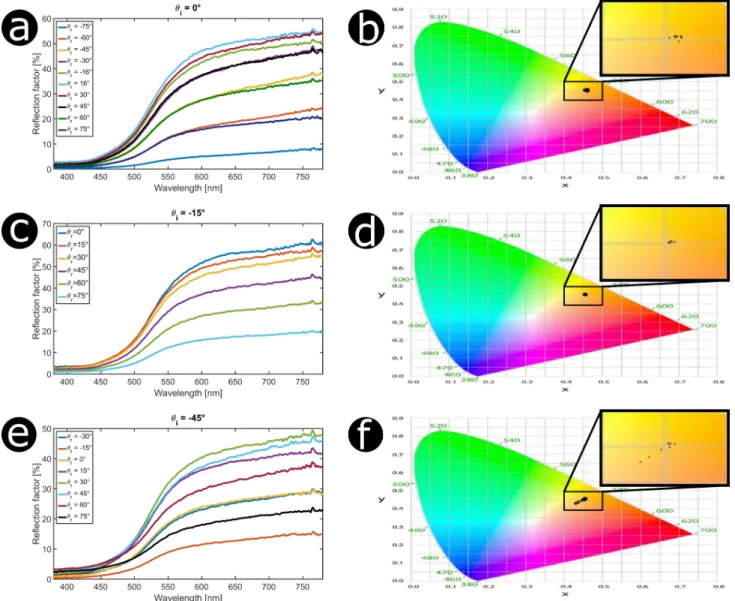

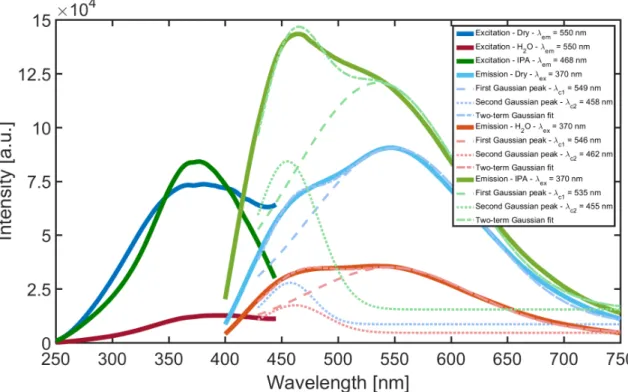
Documents relatifs
A fully-simulated CaSSIS synthetic “true” colour image (RGB) (left) and the CBRC1 image (right) highlight the differences between ferrous- (blue and purple) and ferric-bearing
Using this approach, we reinforce the conclusion that the surface brightness of a star depends on its spectral type and its luminosity class, since our new SBCRs for giants and
Les critères d’évaluation portent, entre autres, sur le degré de réflexivité relatif aux actions restituées et sur la mobilisation des ressources théoriques pertinentes : (i)
The changes occurring in the spectral radiance scattered by spherical grains of different composition and grain size distribution as a function of the phase
We report a new specimen of the extinct procellariiform species Diomedeoides brodkorbi (Aves, Diomedeoididae) from the early Oligocene (Rupelian) of Rheinweiler in
En général, les taux globaux de cas déclarés d’infection à Chlamydia trachomatis chez les femmes sont considérablement plus élevés que les taux correspondants chez les
The soil water contents maintained during the experiment permitted to keep 3 series of 30 poplar cuttings with di fferent levels of water stress, as indicated by predawn leaf water
TRFA measurements were conducted whereby the I VV (t) and I VH (t) fluorescence decays of a series of oligoquinolines labeled at one end with an oligo(phenylene vinylene)
