Effect of plant age and species on population growth of two Rhopalosiphum maidis (Fitch) lineages
Texte intégral
Figure
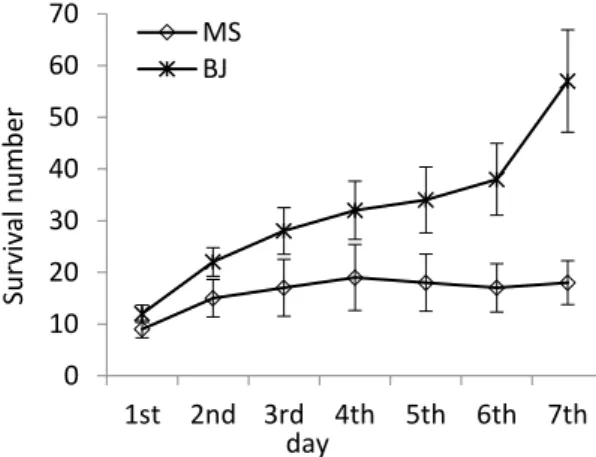
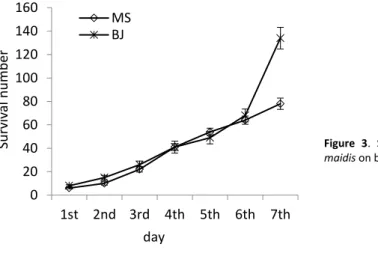
Documents relatifs
C) Individual weighted residuals (IWRES) with respect to time. Population analysis of experimental tumor growth kinetics. A) Visual predictive checks assess goodness-of-fit for
In field experiments aphids were released in the centre of circles of eight potted maize plants. Four plants in each circle were damaged and treated with caterpillar regurgitant
Seasonal changes of leaf specific hydraulic resistance at the whole plant level and for different plant organs, as measured in control (black columns) and infested (dashed
Rough-handling increased fine root leakage and decreased final height and diameter but had no significant effect on survival.. Desiccation had a major effect on the
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
Brown specifically describes the student counselors' responsibil- ities in curriculum orientation, academic adjustment and improvement counseling, survival orientation,
Sont tous les cauchemars assaillant le rêveur Nous sommes les héros de nos propres légendes Fabriquant le passé et ce qui nous transcende Bâillonnés par la peur de la mort qui
To assess the role and genetic diversity of rhizobial bacteria in this agronomic system, we isolated 74 rhizobial strains from root nodules of groundnut, soybean, and cowpea
