Molecular characterization of two monoclonal antibodies specific for the LDL receptor-binding site of human apolipoprotein E
Texte intégral
Figure
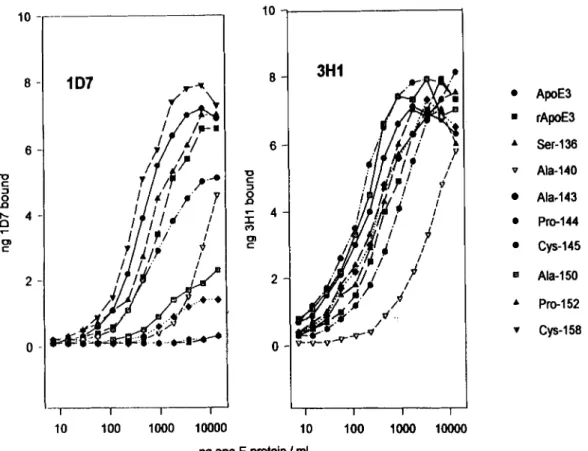
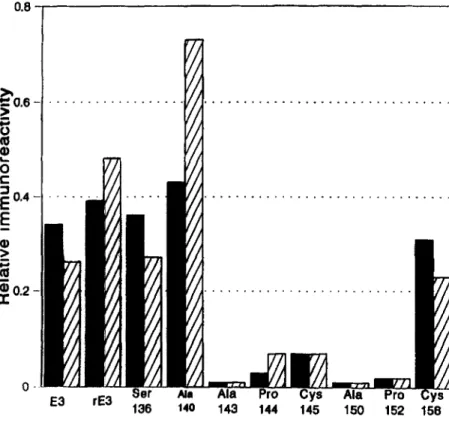
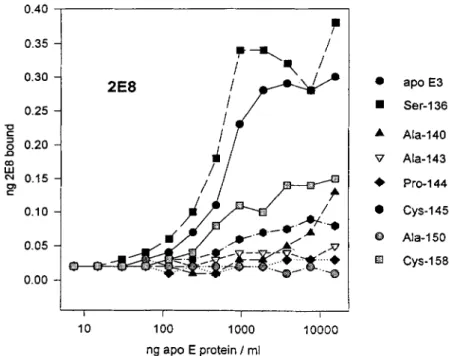
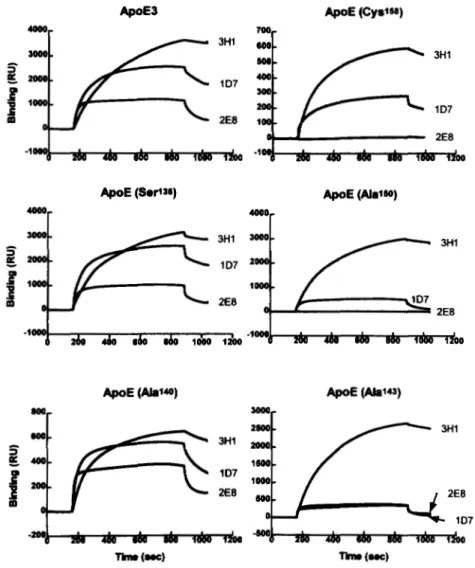
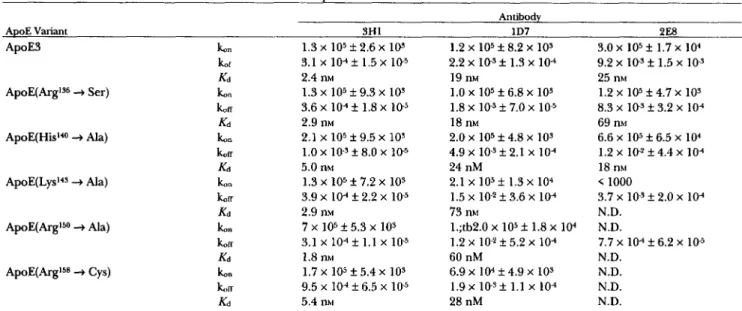


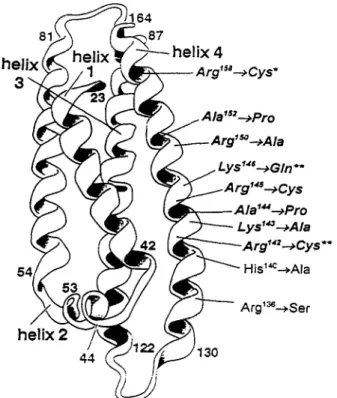
Documents relatifs
However, we believe that the main contribution of this paper is not PairClass itself, but the extension of supervised word pair classification beyond the classification of
ةػػ.. ةمتاخ 126 ةمتاخ ؽافتاب ةحص طرش ونأ ىمع جاكزلا دقع يف دايشلإا ـكقي يف اكفمتخا ـينأ ٌلاإ ،ءايقفلا دنع وطارتشا ىلإ ةيكلاملا بىذك ،دقعلا
Since no specific exercises such as strength training, endur- ance or stretching were imposed during our 5-day multi- modal program, improvements observed in patients with chronic
In this paper, we consider only dynamic graphs in which nodes are anonymous, each of them is infinitely often reachable from any other one, and such that its underlying graph (i.e.,
Nous venons de voir que, pour l’analyse linguistique d’un terme comme noir, il faut éviter l’écueil principal qui consisterait à faire appel aux notions de « désignateur
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
In summary, we designed a new dendritic folate–drug conjugate including a chemical amplifier that enables the receptor-mediated endocytosis and the subsequent enzyme-catalysed
Es gelte also danach zu fragen, welche Normen die Wahrnehmung der Körper- Leiber ‚rahmen‘, inwiefern durch die Markierungen körperbezogener Aspekte Grenzen zwischen Anerkennbarkeit
