Réponses écologiques et évolutives de populations de phytoplancton suite à une acidification
78
0
0
Texte intégral
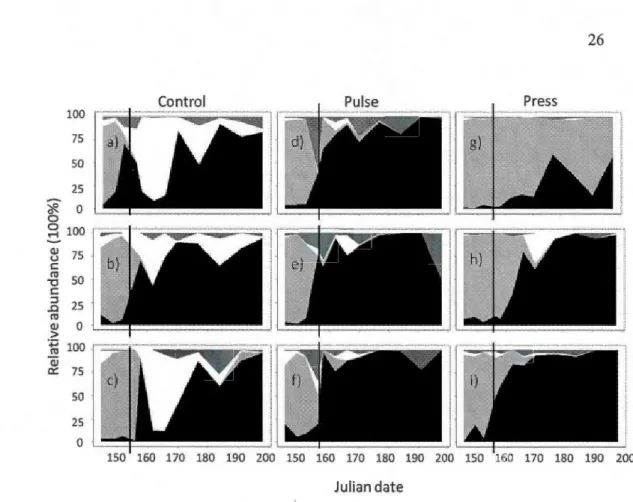
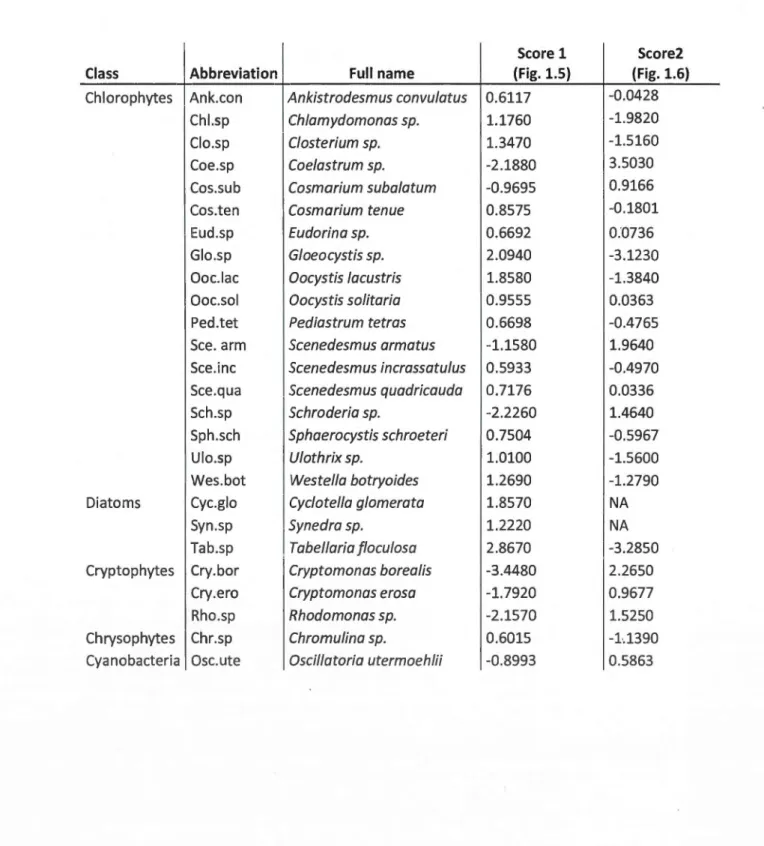
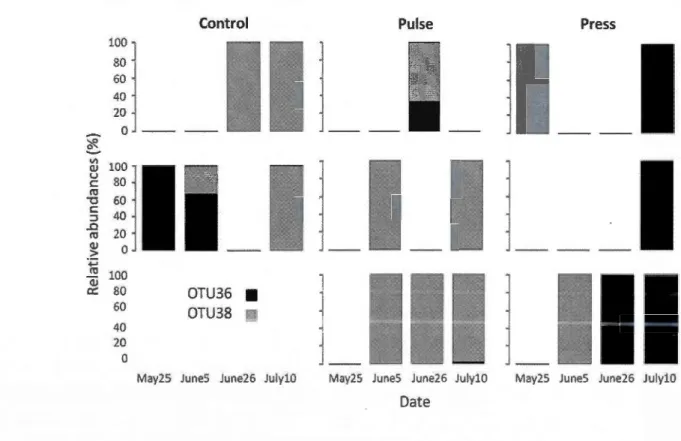
Figure



+7




Documents relatifs