© Lingling Shang, 2019
Basic cultural determinants of recombinant protein yield
in Nicotiana benthamiana used as a transient
expression host for the flu vaccine antigen
hemagglutinin H1
Thèse
Lingling Shang
Doctorat en biologie végétale
Philosophiæ doctor (Ph. D.)
BASIC CULTURAL DETERMINANTS OF RECOMBINANT
PROTEIN YIELD IN NICOTIANA BENTHAMIANA USED AS
A TRANSIENT EXPRESSION HOST FOR THE FLU
VACCINE ANTIGEN HEMAGGLUTININ H1
Thèse
Lingling Shang
Sous la direction de :
Dominique Michaud, directeur de recherche Steeve Pepin, codirecteur de recherche
ii
Résumé
Les plantes sont des hôtes prometteurs pour la production de protéines recombinantes d’intérêt médical et de nombreuses études ont été réalisées au cours des années pour optimiser le taux d’expression des transgènes ou la maturation des protéines en systèmes végétaux. En comparaison, les connaissances demeurent limitées au sujet de l’influence des facteurs environnementaux et des pratiques culturales sur l’expression et le rendement en protéines recombinantes dans les plantes. Les pratiques culturales courantes en serriculture, si elles permettent en général une production importante de biomasse et des rendements élevés en produits horticoles, ne sont pas nécessairement bien adaptés à la production de protéines recombinantes dans un contexte de moléculture. Dans cette étude, nous avons étudié les effets d’un enrichissement en CO2 atmosphérique, d’une forte irradiance sur le
couvert végétal, d’une fertigation riche en ammonium et d’une densité de plantation élevée sur la croissance, le développement et le rendement en protéine recombinante chez l’hôte d’expression Nicotiana benthamiana utilisé pour la production du principe actif d’un vaccin contre la grippe, l’hémagglutinine H1 du virus de l’influenza. En bref, nos données ont montré les effets positifs (1) d’un enrichissement en CO2, d’une forte luminosité, d’un
éclairage intercalaire dans la canopée végétale ou d’une solution nutritive riche en ammonium sur la production de biomasse et le contenu en protéines dans la plante; et (2) d’un éclairage intercalaire et d’une forte densité culturale sur le rendement en protéine H1 par unité de surface en culture. En revanche, le rendement en H1 n’a pas été altéré, ou l’a été négativement, sous de fortes concentrations en CO2 atmosphérique, sous une forte luminosité
au-dessus du couvert végétal ou par une solution nutritive riche en ammonium. En somme, nos données indiquent que les conditions de culture optimales pour la production de produits horticoles en conditions confinées peuvent ne pas être appropriées dans un contexte de moléculture où l’objectif ultime est le rendement en protéine recombinante, non pas la production de biomasse foliaire, la teneur en nutriments ou le rendement en fruits ou en fleurs.
iii
Abstract
Plants are promising hosts for the production of medically-useful recombinant proteins and numerous studies have been done over the years to optimize transgene expression rates and protein maturation processes in plant systems. By comparison, little is still known about the influence of basic environmental factors and cultural practices on the expression and yield of heterologous proteins in plants. Current cultural practices in greenhouse settings, that generally allow for an increased biomass or food/flower product yield, are not necessarily well suited to recombinant protein production in a molecular farming context. In this study,
we investigated the effects of CO2 enrichment, supplemental lighting, ammonium fertigation
and plant culture density on growth, development and recombinant protein yield of the protein expression host Nicotiana benthamiana used to express the flu vaccine antigen influenza virus hemagglutinin H1. In brief, our data showed (1) atmospheric CO2 enrichment,
light irradiance, supplemental LED inter-lighting in the plant canopy and high-ammonium fertigation to enhanced leaf biomass production and endogenous protein content on a plant basis, and (2) LED inter-lighting or elevated plant density to increase recombinant protein yield on a whole-crop area basis. On the other hand, H1 content was not influenced
or negatively affected by CO2 enrichment, high-light irradiance or high-ammonium supply
on a leaf fresh weight basis. Overall, our findings indicate that the optimal cultural practices for the production of horticultural food products or ornementals in controlled environment settings may not be optimal in molecular farming settings, where the ultimate goal is recombinant protein yield and quality, not leaf biomass, nutrient content, fruit yield or flower quality.
iv
TABLE OF CONTENTS
Résumé ... ii Abstract ... iii TABLE OF CONTENTS ... iv LIST OF TABLES ... viLIST OF FIGURES ... viii
ACKNOWLEDGEMENTS | REMERCIEMENTS ... x
FOREWORD | AVANT-PROPOS ... xii
GENERAL INTRODUCTION ... 1
Chapter 1 Literature review, hypotheses and objectives ... 6
1.1 Effects on growth and development ... 8
1.2 Effects on N content ... 13
1.3 Effects on endogenous protein content ... 15
1.4 Effects on recombinant protein yield ... 17
1.5 Working hypotheses and research objectives... 19
Chapter 2 Atmospheric CO2 enrichment and supplemental lighting show differential effects on leaf biomass production and recombinant protein yield in Nicotiana benthamiana transiently expressing the flu vaccine antigen hemagglutinin H1 ... 23
2.1 Résumé ... 24
2.2 Abstract ... 25
2.3 Introduction ... 26
2.4 Materials and Methods ... 29
2.5 Results ... 33
2.6 Discussion ... 41
2.7 Conclusion ... 47
v
Chapter 3 Nutritional ammonium differentially impacts Rubisco content and recombinant protein yield in Nicotiana benthamiana leaves expressing the flu
vaccine antigen hemagglutinin H1 ... 49
3.1 Résumé ... 50
3.2 Abstract ... 51
3.3 Background ... 52
3.4 Materials and Methods ... 54
3.5 Results ... 58
3.6 Discussion ... 69
3.7 Acknowledgements ... 72
3.8 Annexe ... 73
Chapter 4 Effects of CO2 Enrichment, LED Inter-lighting, and High Plant Density on Growth of Nicotiana benthamiana Used as a Host to Express Influenza Virus Hemagglutinin H1 ... 74
4.1 Résumé ... 75
4.2 Abstract ... 76
4.3 Introduction ... 77
4.4 Materials and Methods ... 78
4.5 Results ... 81
4.6 Discussion ... 90
4.7 Acknowledgements ... 97
Conclusions & Perspectives ... 98
vi
LIST OF TABLES
Table 2.1. Effects of atmospheric CO2 enrichment and light integral (PPFD) on the leaf fresh
weight (LFW) of primary stem (P), secondary stem (S) and whole-plant (total, T) leaves from growth chamber-cultivated N. benthamiana plants··· 34
Table 2.2. Effects of atmospheric CO2 enrichment and light integral (PPFD) on leaf area and
leaf mass per area of primary stem (P), secondary stem (S) and whole-plant (total, T) leaves from growth chamber-cultivated N. benthamiana plants··· 35
Table 2.3. Effects of atmospheric CO2 enrichment and light integral (PPFD) on the H1 to
TSP content ratio of primary stem (P), secondary stem (S) and whole-plant (total, T) leaves from growth chamber-cultivated N. benthamiana plants··· 40
Table 3.1. NH4+ and NO3— concentrations in the nutrient solution prior to
agroinfiltration ··· 57
Supplemental Table 1. Nutrient solution used for plants grown at an electrical conductivity
of 2.6 mS/cm (g/ 100L) ···73
Table 4.1. Effects of CO2 concentration, LED inter-lighting, and plant density on plant height,
the leaf/stem fresh weight ratio, and specific fresh weight of primary (P), secondary (S), and total (T) leaves before infiltration··· 81
Table 4.2. Effects of CO2 concentration, LED inter-lighting, and plant density on leaf number
and fresh weight of primary (P), secondary (S), and total (T) leaves per plan ··· 83
Table 4.3. Effects of CO2 concentration, LED inter-lighting, and plant density on leaf number
and fresh weight of primary (P), secondary (S), and total (T) leaves on a whole-crop area basis ··· 84
Table 4.4. Effects of CO2 concentration, LED inter-lighting, and plant density on total soluble
proteins (TSP), protein content per plant, and protein yield per whole-crop surface area
of primary (P), secondary (S), and total (T) leaves at
harvest··· 87
vii
concentration, H1 content per plant, and H1 yield per whole-crop surface area of primary (P), secondary (S), and total (T) leaves at harvest··· 88
viii
LIST OF FIGURES
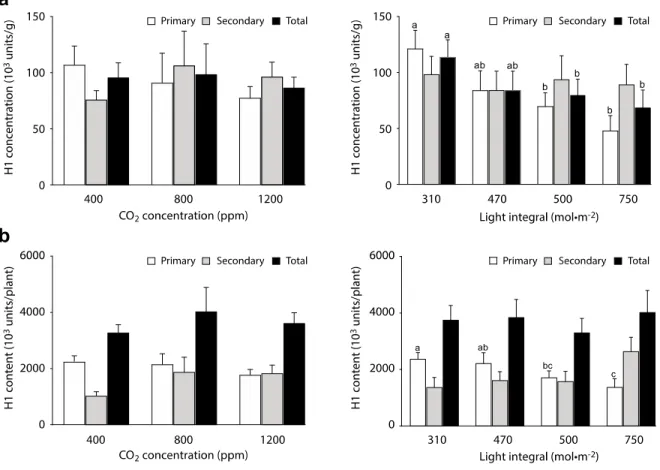
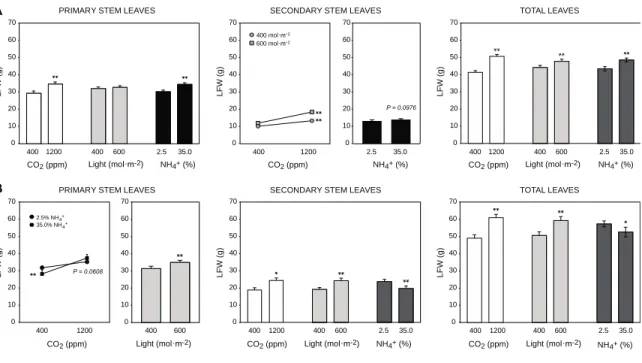
Figure 2.1. Effects of atmospheric CO2 enrichment and light integral (PPFD) on TSP
concentration (A) and content (B) in primary stem, secondary stem and total N.
benthamiana leaves. Each bar is the mean of four independent (replication in time)
values ± se. Different letters on panel B indicate significant differences between light integral treatments at a threshold significance value of 5%, as determined by a protected Fisher’s LSD test. ··· 37
Figure 2.2. Effects of atmospheric CO2 enrichment and light integral (PPFD) on H1
concentration (A) and yield (B) in primary stem, secondary stem and total N.
benthamiana leaves. Each bar is the mean of four independent (replication in time)
values ± se. Different letters indicate significant differences between light integral treatments at a threshold significance value of 5%, as determined by protected Fisher’s LSD tests. ··· 38
Figure 2.3. mRNA transcripts for hemagglutinin H1 in primary stem and secondary stem
leaves of N. benthamiana plants grown under low- (175 μmol·m-2·s-1) or high- (350
μmol·m-2·s-1) PPFD regimes. Each bar is the mean of three independent (time replicate)
values ± se. ··· 41
Figure 3.1. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on the
biomass fresh weight of primary stem, secondary stem and total leaves at the end of the growth period (A) or at harvest following H1 expression (B). Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance thresholds of 5% (*) or 1% (**), as inferred from Tukey’s mean comparison tests··· 60
Figure 3.2. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on total
nitrogen per g LFW in primary stem, secondary stem and total leaves at the end of the growth period before agroinfiltration for H1 expression. Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance thresholds of 5% (*) or 1% (**), as inferred from Tukey’s mean comparison tests··· 62
ix
content per g LFW in primary stem, secondary stem and total leaves at the end of the growth period (A) or at harvest following H1 expression (B). Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance threshold of 5% (*) or 1% (**), as inferred from Tukey’s mean comparison tests··· 63
Figure 3.4. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on H1
hemagglutinin activity per g LFW (A) or on a plant basis (B) for primary stem, secondary stem and total leaves at harvest following H1 expression. Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance thresholds of 5% (*) or 1% (**), as inferred from Tukey’s mean comparison tests··· 64
Figure 3.5. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on Rubisco
large subunit (RbcL) content per g LFW in primary stem, secondary stem and total leaves at the end of the growth period (A) or at harvest following H1 expression (B). Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance thresholds of 5% (*)
or 1% (**), as inferred from Tukey’s mean comparison
tests··· 66
Figure 3.6. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on the TSP to
RbcL ratio in primary stem, secondary stem and total leaves at the end of the growth period (A) or at harvest following H1 expression (B). Each bar is the mean of four independent (replication in time) values ± se. Asterisks (*) indicate significant differences between treatments at a significance threshold of 5%, as inferred from Tukey’s mean comparison tests··· 68
Figure 3.7. Effects of atmospheric CO2 concentration, PPFD and NH4+ supply on the H1
antigen to RbcL ratio in primary stem, secondary stem and total leaves at harvest following H1 expression. Each bar is the mean of four independent (replication in time) values ± se. Asterisks indicate significant differences between treatments at significance thresholds of 5% (*) or 1% (**), as inferred from Tukey’s mean comparison tests··· 69
x
ACKNOWLEDGEMENTS | REMERCIEMENTS
I had the chance, during my Ph.D. program, to be supported by several people. My experience over the last several years let me realize how kind Québécois are, how beautiful Québec City is, and how pleasant it is to work and learn at Laval U. I here take the opportunity to thank these several colleagues, mentors and friends who helped me over the last several years, realizing that this will never be enough.
First and foremost, I wish to express my deep and sincere appreciation to my thesis supervisors. Thank you to Prof. André Gosselin, my main supervisor for the first four years; thank you André for giving me the opportunity to pursue my doctoral studies in Canada and for always being a guide and a great source of motivation and encouragement. Thank you to Prof. Dominique Michaud, my co-supervisor and, more recently, my supervisor; thank you Dominique for your profound knowledge of our research field, for your meticulousness, your patience and your friendliness, that have been of great assistance throughout this last (and toughest) year of my Ph.D. program during which I had to write those manuscripts that now form the core of this thesis. Thank you, finally, to Prof. Steeve Pepin, my co-supervisor; Steeve, you have been for me a great tutor and also a friend; you gave me selfless assistance and support whenever I needed help; you always have been there for me during my ‘confused moments’ or when I had to face those many hurdles that I had to deal with during the course of my work. Thank you, Prof. Gosselin, Prof. Michaud, Prof. Pepin, I am so lucky of having been your student.
I would like to thank my team members for this project, Ms. Linda Gaudreau, Dr. Gilbert Éthier, Ms. Marie-Claire Goulet, Ms. Ann-Catherine Laliberté, Ms. Steffi-Anne Béchard-Dubé, Ms. Jennifer Corriveau Boulay and Ms. Marielle Gagné. Thank you for your valuable
xi
assistance and for always giving me such friendly support. In particular, a big thank you to Linda for providing such instrumental assistance throughout my studies, and to Steffi-Anne for always supporting me and being such a great friend during all these years in Québec City. Thank you, finally, to our colleagues from Medicago inc., for the sustained support and assistance during my experimentations.
Last but not the least, thank you to my dear husband, Dr. Changpeng Qiu who gave me utmost encouragement and support, and to my lovely son Edward Qiu who has been the most wonderful gift during these Ph.D. program years. Thank you to my parents Baoshan Shang and Fuying Dong, to my brother Fengbin Shang, and to my parents in law Yuxing Qiu and Xianglan Xu, who were always there to support me.
Thank you again, and good luck to all of you!
xii
FOREWORD | AVANT-PROPOS
This thesis includes three research manuscripts published, or to be published, in peer-reviewed scientific journals (Chapters 2–4). All chapters, including Chapter 1 (Literature review) and Chapter 5 (Conclusions and perspectives) are in English; a French abstract is also included at the beginning of each research manuscript (Sections 2.0, 3.0 and 4.0, respectively) as suggested by Laval University Faculté des études supérieures et postdoctorales. A consolidated list of references is presented at the end of the document, that includes references cited in all chapters. Several collaborators have contributed to this project, including my supervisor, Dominique Michaud; my co-supervisors, Steeve Pepin and André Gosselin; research professionals Linda Gaudreau, Ann-Catherine Laliberté, Marie-Claire Goulet, Jennifer Corriveau Boulay and Gilbert Éthier; and Michèle Martel, Director Greenhouse Technology at Medicago inc.
CHAPTER 2 (Paper 1) | Shang L, Gaudreau L, Laliberté A-C, Goulet M-C, Gagné M,
Éthier G, Martel M, Michaud D,* Pepin S, Gosselin A (2019) Atmospheric CO2 enrichment
and supplemental lighting show differential effects on leaf biomass and recombinant protein yield in Nicotiana benthamiana transiently expressing the flu vaccine antigen hemagglutinin H1. This manuscript has now been submitted for publication in Molecular Biotechnology. I
contributed to both the experimental design and the experiments, analyzed the data and wrote the first version of the manuscript. L. Gaudreau contributed to the experimental design and to the experiments. A.-C. Laliberté, M.-C. Goulet and M. Gagné contributed to the experiments. G. Éthier and M. Martel contributed to the experimental design. S. Pepin and A. Gosselin contributed to the experimental design and to the first writing of the manuscript. D. Michaud contributed to the experimental design and wrote the final version of the manuscript. All co-authors contributed to the annotation and final proofreading of the manuscript.
xiii
CHAPTER 3 (Paper 2) | Shang L, Gaudreau L, Laliberté A-C, Goulet M-C, Éthier G, Martel
M, Gosselin A, Pepin S, Michaud D* (2019) Nutritional ammonium differentially impacts Rubisco content and recombinant protein yield in Nicotiana benthamiana leaves expressing the flu vaccine antigen hemagglutinin H1. This manuscript has been recently submitted for
publication in Plant Cell Reports. I contributed to the experimental design, performed the experiments, analyzed the data and wrote the first version of the manuscript. L. Gaudreau contributed to the experimental design and to the experiments. A.-C. Laliberté and M.-C. Goulet contributed to the experiments. G. Éthier, M. Martel, S. Pepin and A. Gosselin contributed to the experimental design. D. Michaud contributed to the experimental design and wrote the final version of the manuscript. All co-authors contributed to the annotation and final proofreading of the manuscript.
CHAPTER 4 (Paper 3) | Shang L, Gaudreau L, Martel M, Michaud D,* Pepin S, Gosselin
A (2018) Effects of CO2 enrichment, LED inter-lighting, and high plant density on growth
of Nicotiana benthamiana used as a host to express influenza virus hemagglutinin H1.
Horticulture, Environment and Biotechnology 59, 637–648. This article has been published in October 2018. I contributed to the experimental design and to the experiments, analyzed the data and wrote the first version of the manuscript. L. Gaudreau contributed to the experiments and to the experimental design. M. Martel contributed to the experimental design. S. Pepin and A. Gosselin contributed to the experimental design and to the first writing of the manuscript. D. Michaud contributed to the experimental design and wrote the final version of the manuscript. All co-authors contributed to the annotation and final proofreading of the manuscript.
1
GENERAL INTRODUCTION
The phrase molecular farming (or molecular pharming) was coined in the late 1980’s to describe a new application field in molecular biotechnology dedicated to the production of high-value compounds in transgenic organisms (Bhatia et al., 2015). The term was referring to the heterologous production of clinically-useful proteins in animal hosts, and then in plant hosts, as an alternative to the purification of their native counterparts from natural sources, in a socioeconomic context where the needs in therapeutic proteins were rapidly increasing worldwide and their costs on the market becoming more and more prohibitive (WHO, 2002). Much progress has been achieved over the last three decades in the molecular farming sector, which nowadays mostly refers to the production of clinically- or industrially-valuable recombinant proteins in plant-based systems (Rybicki, 2010; Fischer et al., 2013; Stoger et al., 2014; Tschofen et al, 2016).
Plants present clear advantages as expression hosts for protein pharmaceuticals, including their low upstream production costs, their simple growth requirements, their high potential for large-scale cultivation, and the availability of well-defined schemes for plant biomass harvesting, handling, storage and processing (De Martinis et al., 2016; Xu et al., 2012). Other advantages for molecular farming are the absence of human pathogens in plant tissues, and the intrinsic ability of plant cells to perform protein post-translational modifications– including glycosylation–that match those of mammalian and human cells. A wide array of clinically-valuable proteins have been produced in plant systems over the years, including vaccine antigens (e.g. Lonoce et al., 2016; Tusé et al., 2015; Roychowdhury et al., 2018; Tottey et al., 2018; Vanmarsenille et al., 2018), therapeutic antibodies (e.g. Jutras et al., 2016, Li et al., 2016; Fu et al., 2018; Marusic et al., 2018; Kommineni et al., 2019) and other proteins of clinical interest (e.g. Rattanapisit et al., 2017; Ramirez-Alanis et al., 2018; Silberstein et al., 2018). The commercial potential of plant protein factories has been
2
confirmed recently with the approval of a first plant-made biopharmaceutical for human therapy, the demonstrated feasibility of readily producing therapeutic antibodies in plants during the Ebola virus outbreak some years ago, and the increasing number of plant-made proteins currently undergoing Phase I or Phase II clinical trials (Maxmen, 2012; Merlin et al., 2014; Sack et al., 2015; Lomonossoff and D’Aoust, 2016).
Numerous papers have been published over the last two decades dealing with the optimization of recombinant protein expression in plant systems, most of them focusing on the development of tools and strategies for the improvement of transgene expression, the in
situ stabilization of recombinant proteins or the targeted modulation of protein
post-translational maturation (Gomord et al., 2010; Wen & Zhu, 2015; Jhinjer et al., 2016). In sharp contrast, data are still scarce about the influence of environmental factors and cultural practices on the expression and yield of heterologous proteins in plant systems. A large body of literature has dealt with the impacts of common cultural practices on culture growth and yield in controlled environments, but the best practices in horticultural settings aimed at producing foods or ornementals may not be the best practices in a molecular farming context, where the ultimate target is the yield in recombinant protein, not the production of leaf biomass, flowers or edible foods. From this perspective, our goal in this doctoral project was to improve basic knowledge about the effects of commonly used cultural practices, such as CO2 enrichment, supplemental lighting or ammonium fertigation, on the net yield of a
clinically-useful vaccine antigen, influenza virus hemagglutinin H1 (D’Aoust et al., 2008), produced in leaves of the plant expression host Nicotiana benthamiana, a wild relative of tobacco (Leuzinger et al., 2013).
N. benthmiana has recently become a valuable host for the production of clinically-useful
recombinant proteins (Lomonossoff and D’Aoust, 2016). Used for many years as an experimental model to study plant–pathogen interactions (Goodin et al., 2008), this plant endemic to Australia presents morphological and physiological traits, such as a rapid growth
3
rate, a large leaf biomass index and a natural ability to express heterologous gene sequences, that make it an interesting host for the transient expression of several recombinant proteins (Bally et al., 2018). Heterologous expression procedures involving N. benthamiana generally rely on the vacuum infiltration of leaf tissue with Agrobacterium tumefaciens cells harboring a transgene for the protein of interest (Kapila et al., 1997). Efficient procedures have been developed to express heterologous genes in N. benthamiana, that now allow for the production of milligrams of recombinant protein in a short period of time using only a handful of plants (Leuzinger et al., 2013; Norkunas et al., 2018).
Our goal in this project was to assess the effects of atmospheric CO2 concentration, light
irradiance, mineral nitrogen (N) nutrition and plant cultural density on basic growth parameters, endogenous protein content and recombinant protein (H1) yield of N.
benthamiana leaves set up to express the viral antigen under the control of a constitutive
promoter. These basic cultural factors, given their strong influence on plant growth and development, have been the object of intense research over the years that led to the development of well adapted cultural practices in horticulture production settings. For
instance, atmospheric CO2 enrichment and supplemental lighting have now become common
cultural practices in greenhouse settings that, if managed properly, allow for a significant increase of biomass production and fruit (or flower) yield. Here, we assessed the potential of these practices from a molecular farming perspective considering, on the one side, the well-established importance of leaf biomass on recombinant protein yields in N. benthamiana (Matsuda et al., 2018) but, on the other side, the lack of data linking biomass production, endogenous protein biosynthesis and foreign protein accumulation with cultural conditions normally adopted to promote basic plant growth and energy deposition in sink tissues (e.g. fruits) or storage compounds (including Rubisco in leaves).
Simply put, recombinant protein yield in N. benthamiana protein production settings is determined by three main factors: (1) the specific amount of recombinant protein in
4
agroinfiltrated leaf tissue (in units [or mg]·g fresh weight-1); (2) the leaf biomass per plant
prior to infiltration and/or at the end of the protein expression period (in g·plant-1 or g·m-2); and (3) the number of plants per m2 of growing area before agroinfiltration (Fujiuchi et al., 2016; Goulet et al., 2019). A fourth, complementary factor is the amount of ribulose 1,5-biphosphate carboxylase oxygenase (Rubisco) in leaf tissue, as this protein not only is an indicator of plant growth (Erb and Zarzucki, 2018) but also a strong sink for amino acids storage under favorable nutrient conditions (Millard, 1988) and a major endogenous contaminant upon recombinant protein recovery and purification (Buyel et al., 2015; Robert et al., 2015). Taking these four yield-driving factors into consideration, the present project involved three main experiments exploring, together, the effects of atmospheric CO2
enrichment, supplemental lighting, ammonium nutrition and plant culture density on leaf biomass production, H1 antigen titer following heterologous expression, Rubisco content before and after H1 expression, and net H1 yield under different plant culture densities in an experimental greenhouse production setting.
The core of the thesis includes an introductory literature review chapter to describe the project’s research problem, the working hypotheses and the specific objectives pursued; one published scientific article in a peer-reviewed journal and two others recently submitted to other journals; and a concluding chapter that summarizes the main findings and their meaning considering the working hypotheses formulated a priori. Chapter 1 draws an overview of current knowledge about the impacts of basic environmental factors and common cultural practices on growth and development of plants produced under controlled environments, followed by an enumeration of the project’s working hypotheses and research objectives.
Chapter 2 (Article 1) and Chapter 3 (Article 2) present the results of Experiments 1 and 2
comparing the leaf biomass, endogenous protein content, Rubisco content and H1 antigen
yield of plants produced in growth chambers under different atmospheric CO2 concentrations,
light intensities and nitrogen nutritional regimes. Chapter 4 (Article 3) presents the results
5
and plant culture density on H1 yield, leaf soluble protein content and different growth parameters of plants produced in a glasshouse setting. Chapter 5 concludes the thesis with a short summary on the project’s main findings, a post-experiments assessment of the working hypotheses and some new ideas and questions for future work on cultural practices in N. benthamiana heterologous protein expression settings.
6
Chapter 1
Literature review, hypotheses and objectives
About the influence of basic cultural conditions on plant growth,
nitrogen uptake and protein content
7
The influence of environmental factors on plant growth and development has been thoroughly assessed over the years, in relation with their resulting impacts on yield and quality in crop production settings. More specifically, numerous studies have considered the
impacts of cultural practices such as atmospheric CO2 enrichment and supplemental lighting
in confined environments on the biomass, nutrient content, fruit yield and/or flower quality of horticultural crops. From an R&D standpoint, the effectiveness of a given cultural practice for crop yield improvement is first based on a thorough understanding of sink-source relationships in planta, followed by the successful transfer of this basic knowledge in a horticultural context. Because the ultimate goal of molecular farming is not to maximize biomass or fruit harvest but to produce exogenous proteins that are not expressed naturally in the plant, the actual usefulness of common cultural practices in these systems remains to be assessed as several questions of practical relevance remain unanswered. Here is one such question, among others: are basic cultural practices known to promote endogenous protein accumulation in leaves, such as a nutrient solution containing a significant concentration of
ammonium (NH4+), also promoting the accumulation of a foreign protein?
We aimed in this project at assessing the impacts of commonly used cultural practices like atmospheric CO2 enrichment, supplemental lighting and varying-NH4+ nutritional supply on
growth, protein content and recombinant protein yield of N. benthamiana used as a transient expression host for the influenza virus vaccine antigen hemagglutinin H1 (D’Aoust et al., 2010). A series of experiments were conducted toward this goal, as described in Chapters
2–4. A short literature review is here presented as a baseline for further discussion, about the
impacts of atmospheric CO2 concentration, light irradiance and nitrogen (N) supply on
growth, development, N uptake and protein content of plants cultivated in contained environments.
8
1.1
Effects on growth and development
1.1.1 Atmospheric CO2 concentration
Atmospheric CO2 concentration is a major yield determinant in controlled environment
production settings given its direct impact on the carboxylase/oxygenase activity balance of
ribulose 1,5-bisphosphate carboxylase oxygenase (Rubisco), a key enzyme for CO2 fixation
in higher plants (Drake et al., 1997). In practice, CO2 enrichment increases the net
carboxylation rate of Rubisco and promotes biomass production. Considering 15 years of
CO2 enrichment experiments, Ainsworth and Long (2005) estimated the light-saturated leaf
photosynthetic rate to be increased by 31% on average in C3 crops, associated with increased
branching rates, leaf numbers and above-ground dry matter. Likewise, Lindhout and Pet (1990) inferred an average growth rate enhanced by a factor of 2.3 compared to control plants for different tomato genotypes grown under 750 ppm CO2, similar to Tamoi et al. (2011)
reporting increased lateral shoot outgrowth, leaf numbers and leaf biomass upon CO2
enrichment for transgenic tobacco lines ectopically expressing the sugar-metabolizing enzyme fructose-1,6-bisphosphatase.
The growth-promoting effect of CO2 enrichment, however, is usually not maintained over
long periods of time. In most species, the initial stimulation of photosynthesis under
high-CO2 environments is followed after two or three weeks by the gradual establishment of an
‘acclimation state’ that triggers photosynthesis suppression in green tissues (Mauney et al., 1978; Bowes, 1993). For instance, cucumber plants kept in growth chambers under 1,000
ppm CO2 were 83% heavier than control plants grown under 350 ppm CO2 after 16 days, but
only 11% heavier than, or the same weight as, control plants after 36 and 60 days, respectively (Peet et al., 1986). The same phenomenon was observed for several other plants
9
including Solanaceae species, such as tomato plants under 900 ppm CO2 that showed relative
growth rates significantly increased after two weeks but lower than those of control plants after ten weeks (Yelle et al., 1990).
Plant acclimation to elevated CO2 concentrations is the result of a negative feedback response
consequent to an imbalance in the supply and demand for carbohydrates in sink and source
organs (Arp, 1991). Under this model, accumulated starch in high CO2-grown plants (Nie et
al., 1995; De Souza et al, 2008; Aranjuelo et al., 2011) causes an alteration of sink-source relationships in the plant that is eventually counterbalanced by a reduced photosynthetic rate (Usuda and Shimogawara, 1998; Usuda, 2006). The hypothesis of an altered pattern of sink-source relationships triggering CO2 acclimation in C3 crops is supported by the observation
that photosynthesis suppression is frequent in pot-grown plants but rarely reported for soil-grown plants (Arp and Drake, 1991). Given a limited volume of substrate, pot-soil-grown plants present poorly developed root systems inefficient in attracting leaf-produced photosynthates (Heret and Joliffe, 1985). Upon transfer to ambient CO2 conditions, photosynthesis in these
plants is fully restored within a few days as the stored sugars are gradually hydrolyzed and the initial sink-source relationships reinstalled at the whole-plant scale (Hofstra & Hesketh, 1975; Sasek et al., 1985).
No information is yet available in the literature about the effects of CO2 enrichment on N.
benthamiana growth in controlled environment settings, or on the relative balance between
endogenous (host) and heterologous (foreign) protein expression in N. benthamiana leaves following agroinfiltration. Likewise, no data are available about how atmospheric CO2
concentration influences the morphology of this plant. Assessing the data of 120 primary research studies, Ainsworth and Long (2005) reported the specific leaf area of high CO2
10
significant increase of specific leaf weight (SLW). Indeed, high CO2-grown plants generally
tend to be sturdy, and their leaves to be thick (Tremblay and Gosselin, 1998; Yin, 2002), which might eventually prove of practical value to reduce yield losses due to leaf damage. At the end of the growth period, N. benthamiana plants are placed upside down and their above-ground parts are submerged in an Agrobacterium suspension culture for vacuum infiltration (Leuzinger et al., 2013). Under these conditions, the plants suffer mechanical damage and some fragile petioles, stems and leaves are inevitably lost prior to the protein expression phase. Using robust, sturdier plants at this stage of the procedure could have a positive impact on overall protein yield as expressed on a culture area basis, especially in large-scale production settings where automatic, multi-plant handling operations are implemented.
1.1.2 Light irradiance
While atmospheric CO2 is the primary limiting factor of photosynthesis in C3 plants, light
irradiance is the driving force of this process as it directly determines the number of photons intercepted and absorbed in the plant canopy. In practice, the amount of light effectively reaching the leaves is closely determined by plant density in the culture canopy, as defined as the number of plants per growing area unit. Increasing plant density generally has a direct positive impact on total yield, up to a certain level above which a coordinated response to low-light conditions is induced in the plant. This adaptive response, referred to as the shade avoidance syndrome, is initiated by phytochrome photoreceptors in leaves that perceive a reduction of light quantity when the plant is shaded by neighboring plants (Roig-Villanova and Martínez-García, 2016; Ballaré and Pierik, 2017). Typical symptoms of the shade avoidance syndrome include an altered morphology of the plant and a reduced total biomass (Suzuki et al., 2009; Ballaré and Pierik, 2017; Khan et al., 2017). Pierik et al. (2004) reported
11
for instance the stem length/shoot dry weight to be significantly increased in tobacco at elevated plant culture densities, like Seif et al. (2016) describing taller snap bean plants with a reduced leaf area and less dry weight per plant under similar conditions.
Light intensity also has a direct influence on plant morphology. For instance, Schoch et al. (1990) reported the stem diameter of tomato plants during the vegetative growth period to be proportional to the overall amount of light received by the plant, similar to Fan et al. (2013) reporting high photosynthetic photon flux density (PPFD) regimes to decrease the specific leaf area of young tomato plants. On their side, Fu et al. (2017) reported that lettuce plants were stocky under high light intensity but slender under low intensity, in agreement with
Tremblay and Gosselin (1998) who showed that high PPFD (and CO2 enrichment) induce(s)
sturdier, more compact plants in transplant production. These studies, like several others, supported overall the well-established idea that plants grown under high-PPFD conditions
(like those grown under elevated atmospheric CO2 levels) are sturdier than plants provided
with lower amounts of light (or CO2).
Little is known about the light requirements of a desert plant like N. benthamiana in controlled environment (e.g. greenhouse) settings. Likewise, no information is available about possible synergistic effects between supplemental lighting and CO2 enrichment on
plants used for recombinant protein production. Dorais et al. (2017) reported that the
efficiency of supplemental lighting is increased under high-CO2 conditions in
greenhouse-grown fruiting vegetable productions, similar to Pérez-López et al. (2015) showing that high light intensity over the plant canopy could increase biomass production in lettuce under elevated CO2 concentrations. At present, a general lesson from the scientific literature is that
basic environmental factors like CO2 concentration and light irradiance conditions should be
12
environment settings, whether the plants are grown for agricultural purposes or as expression hosts for medically valuable recombinant proteins.
1.1.3 N regime
N is a key constituent of many important compounds in plants including amino acids, proteins, nucleic acids, chlorophyll, hormones and a wide array of defense chemicals. Found under different forms –organic or inorganic– in the soil substrate, this element is required in large quantities by the plant and represents a major growth-limiting factor in most ecosystems. The roots of higher plants mainly absorb N as nitrate (NO3–) and ammonium (NH4+), that then
undergo metabolic assimilation through their integration into amino acids. NH4+ cations
present a reduced form compatible with the metabolism and are assimilated directly, mostly in root tissues shortly after their absorption by hairy roots. NO3– anions, often the most
abundant in the soil solution, are readily translocated in the xylem towards above-ground organs and stored in the cell vacuoles of stems and leaves, before their reduction to NH4+ by
nitrate and nitrite reductases and their subsequent assimilation in the general metabolism.
Despite higher energy costs for assimilation (Huppe and Turpin, 1994), most plants prefer NO3– over NH4+ and show symptoms of toxicity at elevated NH4+:NO3– ratios in the nutrient
solution. For instance, Lu et al. (2005), and Zou et al. (2005), reported low growth rates for tobacco plants assigned an NH4+-only regime compared to higher growth rates for plants
provided with NH4+:NO3– ratios of 50% or 0% (NO3–-only regime). Likewise, Fallovo et al.
(2009) observed the shoot biomass, leaf area and leaf number of two Brassica species to be greater under an NO3–-only diet than under a 50% NH4+ or an NH4+-only diet, similar to
Karaivazoglou et al. (2007) reporting early growth delays for tobacco plants given only NH4+.
13
certain threshold, notably involving toxic effects of free ammonia in planta, acidification of the substrate solution, carbohydrate limitation due to excessive consumption of soluble sugars, and interfering effects limiting the absorption of K+, Ca2+ or Mg2+ (Esteban et al., 2016). Low intracellular concentrations of NO3– leading to osmotic imbalance and water
deficit as a result of reduced water uptake have also been proposed as possible growth-limiting factors at high NH4+ concentrations. Other studies have proposed a link between
NH4+ uptake and low concentrations of growth hormones, such as cytokinins, that promote
cell division and organ morphogenesis (Lu et al., 2009).
Even though some plants show a net preference for NH4+ (Britto and Kronzucker, 2002;
Tsabarducas et al., 2017), a quick look at the literature readily confirms the positive impact
of NO3– uptake on the development of most plants (Coruzzi and Bush, 2001) and the growth
benefits for several plants of absorbing both NO3– and NH4+ (Britto and Krunzucker, 2002).
Several authors have described closely integrated physiological effects for these two nutrients in planta (Hachiya et al., 2017) and the positive effects of a well-balanced NH4+:NO3– ratio in the nutrient solution on growth and total biomass of horticultural crops
in hydroponic production settings have been described in several studies (Marschner, 2002; Sonneveld, 2002; Savvas et al., 2006; Lu et al., 2009; Hu et al., 2017; Liu et al., 2017; Wang et al., 2019). Little is known about the relative effects of NH4+ and NO3– on N. benthamiana
growth but a positive impact of NH4+ at moderate relative concentrations appears plausible
given the published literature on Solanaceae species, including tobacco.
1.2
Effects on N content
CO2 CONCENTRATION |Cultural conditions may have a significant impact on total N content
14
acids and proteins, whether endogenous or recombinant. In particular, CO2 enrichment
generally has a negative impact on total N concentration in planta (Ainsworth and Long, 2005), that notably results from a dilution effect in plants fixing more CO2 for sugar
biosynthesis, a higher N use efficiency index and/or a decreased transpiration rate per plant (Berntson, 1994; McDonald et al., 2002; Leakey et al., 2009; Pang et al., 2006; Taub and Wang, 2008). After systematically reviewing 75 primary research studies, Cotrufo et al.
(1998) estimated high atmospheric CO2 concentrations to reduce total N concentration by 14%
on average in above-ground tissues. Similarly, Han et al. (2015) found the N concentrationof winter wheat at different developmental stages to decrease by 3 to 6% under high CO2
conditions in a free-air CO2 enrichment experimental setup, in line with Geiger et al. (1999)
reporting reduced concentrations of NH4+, NO3– and total amino acids in tobacco leaves
under similar conditions.
N REGIME |Similar to CO2 concentration, NH4+:NO3– ratios in nutrient solutions may have a
strong impact on N concentration in the plant. Walch-Liu et al. (2001) reported for instance that total N concentration in shoots of tobacco was greater in plants provided an NH4+:NO3–
ratio of 50% than in plants given only NO3–. Likewise, Zou et al. (2005) showed N
concentration in tobacco leaves to be higher in plants fertilized with NH4NO3 than in plants
given NH4+ or NO3– only, in line with Liu et al. (2017) reporting an incremental increase of
N concentration in leaves of tomato seedlings given this element at increasing NH4+:NO3–
ratios in the nutrient solution. No study has yet documented the relative effects of NO3– and
NH4+ on total N content in N. benthamiana but the measured effects in tobacco and tomato,
similar to those also observed in other Solanaceae including cucumber (Heuer, 1991), pepper (Bar-Tal et al., 2001), eggplant (Savvas et al., 2010) and stamnagathi (Chatzigianni et al., 2018), suggest an eventual positive effect of the cationic nutrient in this plant, at least up to a certain concentration threshold.
15
1.3
Effects on endogenous protein content
CO2 CONCENTRATION | Cultural conditions that influence total N concentration in planta may
also have an impact on endogenous protein content. In particular, high-CO2 conditions, via
their positive effects on leaf biomass and their negative effect on N concentration, positively influence the C/N balance in planta, with a resulting negative impact on total soluble proteins (TSP) content. For instance, Xu et al. (2014) reported reduced TSP levels on an LFW basis in Gerbera jamesonii leaves upon CO2 enrichment, in line with Barbehenne et al. (2004)
reporting TSP levels reduced by 12% or 6% under high-CO2 environments in C3 and C4
grasses, respectively. Similarly, Högy et al. (2009) found the TSP content in wheat grain to
decrease by more than 7% upon CO2 enrichment, in accordance with a large-scale,
multi-species meta-analysis by Taub et al. (2008) indicating a systematic decrease of TSP content in plants at elevated atmospheric CO2 concentrations.
Despite an obvious trend downwards, the net impact of CO2 enrichment on TSP content is
highly variable depending on the plant species or on free N availability in plant tissues or the nutritive solution. For instance, Vu et al. (1989) reported TSP concentration in leaves to be
increased by 31% in soybean (Glycine max [L.] Merr.) plants grown under 800 ppm CO2,
likely allowed by sufficient N metabolic resources in this N-fixating species. By comparison, Geiger et al. (1999) showed CO2 enrichment to reduce the leaf TSP content in tobacco
provided low (2 mM KNO3) or intermediate (3 mM NH4NO3+ 6 mM KNO3) concentrations
of mineral N. This expected penalty on protein content upon CO2 enrichment was avoided in
plants provided N at 10 mM or 20 mM in the nutrient solution, again supporting the idea of an N resources shortage in plants overproducing carbon skeletons for biomass production under high-CO2 conditions.
16
No data are yet available about the influence of CO2 concentration on TSP content –and
Rubisco content– in N. benthamiana leaves. As a general rule, Rubisco content is reduced in leaves and other green tissues upon CO2 enrichment (Bloom et al., 2002; Drake et al., 1997;
Gamage et al., 2018; Nakano et al., 1997). High-CO2 conditions, by their strong positive
effect on photosynthesis and carbohydrate biosynthesis in leaf cells, induce the accumulation of non-structural carbohydrates and a parallel decline of Rubisco subunits in the chloroplast stroma (Seneweera et al., 2011). As proposed by several authors, these non-structural carbohydrates would hinder the biosynthesis of Rubisco large (RbcL) and small (RbcS) subunits by repressing the transcription of their DNA coding sequences in both the chloroplast and the nucleus (Moore et al., 1999; Takatani et al., 2014; Vicente et al., 2015).
This downregulating effect of CO2 enrichment on Rubisco content could prove of eventual
value in a molecular farming context, considering that this enzyme not only is an indicator of plant growth (Erb and Zarzycki, 2018) but also a problematic endogenous contaminant upon recombinant protein recovery from source tissues (Buyel et al., 2015; Robert et al., 2015).
N REGIME | Similar to CO2 concentration, the N regime may have a significant impact on
endogenous TSP content. In general, high-NH4+ regimes increase protein content in leaves
(Akl et al., 2003; Horchani et al., 2010; Borgognone et al., 2013), in accordance with the low energy costs required for NH4+ assimilation (Huppe and Turpin, 1994) and the
well-established involvement of endogenous proteins, notably Rubisco, in the storage of N resources (Millard, 1988). For instance, Horchani et al. (2010) measured TSP concentrations in tomato plants given 10 mM Ca(NO3)2 in the nutritive solution lower than in plants given
10 mM (NH4)2SO4. Similarly, Raab and Terry (1994) found TSP levels in sugar beet (Beta
vulgaris L. cv F58-554H1) to be higher in plants given an NH4+-only diet (NH4)2SO4 than in
17
tomato (Borgognone et al., 2013). No information is yet available, however, about the relative effects of NH4+ and NO3– on endogenous TSP content and Rubisco concentration in
N. benthamiana leaves.
1.4
Effects on recombinant protein yield
Compared to leaf biomass production, TSP content or fruit yield in horticultural production settings, little attention has been paid until now to the influence of cultural factors like CO2
concentration, N nutrient supply, supplemental lighting or plant culture density on recombinant protein yields in plant protein bio-factories. Nevertheless, a number of papers dealing with those questions clearly suggested the relevance of additional experimental work where these plants are first considered as heterologous protein bioreactors, not as biomass producers. In particular, studies have assessed the effects of light irradiance on recombinant protein production but no general conclusion could be reached as per the net impacts of this cultural factor on yield and the actual relevance of supplemental lighting in molecular farming. For instance, Stevens et al. (2000) reported a positive relationship between an increased PPFD and mammalian antibody accumulation in transgenic tobacco lines, while Matsuda et al. (2012) observed no effect for the same cultural practice with N. benthamiana leaves transiently expressing the H1 vaccine antigen. Likewise, Gaudreau et al. (2015) have shown that increasing light integral during the growth period before agroinfiltration may to some extent have a positive impact on H1 yield in N. benthamiana leaves, but that it may have a negative impact above a certain threshold.
Complementary studies on supplemental lighting will be warranted in coming years to further document the impacts of light irradiance on recombinant protein yield, notably in combination with basic cultural factors like atmospheric CO2 concentration, plant culture
18
density and nutrient solution composition (Colgan et al., 2010; Fujiuchi et al., 2014; Okamura et al., 2014). Studies will also be warranted to evaluate the impacts of cultural conditions taking the host plant general morphology into account, given the strong impact of leaf physiological age on recombinant protein distribution patterns (Stevens et al., 2000; Robert et al., 2013; Béchard Dubé, 2015; Jutras et al., 2016) and the possible differential effects of these conditions on different parts of the plant (Goulet al., 2019). In this regard, Colgan et al. (2010) reported that a high plant density may have no impact on recombinant protein yield in young leaves of transgenic tobacco lines but a significant negative impact in older (bottom) leaves, regardless of the light irradiance. Similarly, Goulet et al. (2019) recently reported that the net effects of cultural practices such as supplemental lighting or treatment with growth-promoting hormones highly depend on leaf position (and leaf age) in N. benthamiana transiently expressing the H1 vaccine antigen.
Interesting studies have been published in recent years assessing the impacts of cultural conditions on recombinant protein yields but numerous questions still remain unanswered at this point. In this context, my goal for this project was to address some of these questions, related in particular to the combined effects of supplemental lighting, atmospheric CO2
enrichment, varying nutrientNH4+ supply in combination with NO3– supply and/or plant
culture density on biomass production, endogenous TSP content and recombinant protein yield in the N. benthamiana transient expression host. More specifically, I have addressed the following three questions:
. QUESTION 1 Do (eventual) positive effects of CO2 enrichment and supplemental
lighting on biomass production during the growth period promote recombinant protein yield in N. benthamiana leaves at the end of the protein expression phase?
19
. QUESTION 2 Do (eventual) positive effects of nutritive NH4+ on biomass production
and endogenous protein content during the growth period promote recombinant protein yield in N. benthamiana leaves at the end of the protein expression phase?
. QUESTION 3 Do plant density and within-canopy supplemental lighting in an N.
benthamiana protein production setting during the growth period influence recombinant
protein yield on a plant culture area basis at the end of the protein expression phase?
1.5
Working hypotheses and research objectives
Three working hypotheses were tested, and three general research objectives pursued for each of them, to address the above-formulated three questions. N. benthamiana was used as host plant model (Lomonossoff and D’Aoust, 2016), and the H1 vaccine antigen as a recombinant protein model of medical value (D’Aoust et al., 2008), for all the experiments.
Chapter 2 (Paper 1) addresses my first hypothesis (Working hypothesis No. 1); Chapter 3
(Paper 2) my second hypothesis (Working hypothesis 2); and Chapter 4 (Paper 3) my third hypothesis (Working hypothesis 3).
FIRST WORKING HYPOTHESIS
Considering (1) many studies that conclude that CO2 enrichment and high light irradiance
have positive impacts on the leaf biomass and robustness of C3 plants (Ainsworth and Long,
2005); and (2) the relationship established by some authors between total leaf biomass and recombinant protein yield in N. benthamiana (reviewed in Fujiuchi et al., 2016), we formulated the following first hypothesis for the project :
20
Working hypothesis No. 1 | Atmospheric CO2 enrichment and high light irradiance
increase H1 yield in N. benthamiana by a positive effect on leaf biomass production.
The following three research objectives were pursued to test this hypothesis:
. OBJECTIVE 1.1 To investigate the effects of CO2 enrichment and high light
irradiance on N. benthamiana main (primary) stem, axillary (secondary) stem and total (primary stem + secondary stem) leaf biomass.
. OBJECTIVE 1.2 To characterize morphological changes on N. benthamiana
plants upon atmospheric CO2 enrichment and/or under high irradiance conditions, as inferred
from measurable changes of specific leaf weight or leaf biomass distribution among primary and secondary stems.
. OBJECTIVE 1.3 To determine the relative effects of CO2 enrichment and high
light irradiance on H1 antigen yield in N. benthamiana primary stem and secondary stem leaves.
SECOND WORKING HYPOTHESIS
Considering (1) the positive impact of a well-balanced NH4+:NO3– ratio on growth and total
biomass of horticultural crops in hydroponic production settings (Marschner, 2002; Sonneveld, 2002; Savvas et al., 2006; Lu et al., 2009; Hu et al., 2017; Liu et al., 2017; Wang et al., 2019); (2) the positive impact of a high-NH4+ supply on TSP content in several plants
21
atmospheric CO2 concentration (Li et al., 2007; Bloom et al., 2012; del Amor et al., 2015;
Vega-Mas et al., 2015; Rubio-Asensio and Bloom, 2016) and light irradiance (Fallovo et al., 2009; Setién et al., 2013; Alencar et al., 2019) on N assimilation and NH4+:NO3– interactions
in planta, we formulated this second hypothesis for the project :
Working hypothesis No. 2 | High ammonium supply in the nutrient solution
promotes H1 yield in N. benthamiana via a positive effect on leaf biomass and protein content.
The following three research objectives were pursued to test this hypothesis:
. OBJECTIVE 2.1 To investigate the effects of low- and high-NH4+ nutritive
regimes on the biomass, total N concentration, TSP content and H1 yield of N. benthamiana primary stem and secondary stem leaves.
. OBJECTIVE 2.2 To investigate the effects of low- and high-NH4+ nutritive
regimes on Rubisco concentration in primary stem and secondary stem leaves, in relation with their corresponding impacts on TSP content and H1 yield.
. OBJECTIVE 2.3 To detect eventual interactions between atmospheric CO2
concentration, light irradiance and NH4+ supply on H1 antigen yield in N. benthamiana
22 THIRD WORKING HYPOTHESIS
Considering (1) the positive effects of within-canopy inter-lighting on light use efficiency and plant growth in greenhouse settings (Koivisto & Hovi-Pekkanen, 2008; Pepin et al., 2013; Guo et al., 2016); and (2) increasing empirical evidence linking high plant density with high recombinant protein yields per unit of culture area in molecular farming settings (Fujiuchi et al., 2016), we formulated this third hypothesis for the project:
Working hypothesis No. 3 | Plant density and within-canopy supplemental lighting
during the growth period positively influence H1 yield on a plant culture area basis.
The following three research objectives were pursued to test this hypothesis:
. OBJECTIVE 3.1 To investigate the effects of high-density planting and
supplemental inter-lighting on the biomass and H1 antigen yield of N. benthamiana primary stem and secondary stem leaves.
. OBJECTIVE 3.2 To evaluate the net impacts of these same two cultural practices on total H1 yield on a culture area basis in an N. benthamiana greenhouse production setting.
. OBJECTIVE 3.3 To detect eventual interactions between supplemental inter-lighting, high plant density and CO2 enrichment used as complement to the first two cultural
23
Chapter 2
Atmospheric CO
2enrichment and supplemental lighting show
differential effects on leaf biomass production and recombinant
protein yield in Nicotiana benthamiana transiently expressing
the flu vaccine antigen hemagglutinin H1
Cet article, co-signé avec Linda Gaudreau, Ann-Catherine Laliberté, Marie-Claire Goulet, Marielle Gagné, Michèle Martel, Dominique Michaud, Steeve Pepin et André Gosselin, a été soumis récemment pour publication dans le périodique scientifique Molecular Biotechnology.
24
2.1 Résumé
Peu de données sont disponibles concernant l’influence des conditions culturales sur le rendement en protéine recombinante dans les plantes utilisées comme hôtes d’expression pour la production de protéines d’intérêt médical. Nous avons ici étudié les effets de la teneur
en CO2 atmosphérique et de la densité de flux photoniques photosynthétiques appliquée au
couvert végétal sur la croissance et le développement de l’hôte végétal Nicotiana
benthamiana utilisé pour la production d’un antigène antigrippal, l’hémagglutinine H1 du
virus de l’influenza. En bref, nos données ont montré qu’un enrichissement en CO2
atmosphérique et une forte quantité de lumière dans la chambre permettaient d’augmenter la biomasse foliaire et la masse spécifique des feuilles pendant la phase de croissance mais conduisaient en revanche à une baisse du contenu en H1 par unité de biomasse foliaire fraîche, en comparaison aux protéines solubles totales. Exprimées pour l’ensemble de la plante, nos données ont montré une augmentation du contenu en protéines solubles sous forte intensité lumineuse mais un effet négligeable sur le rendement total en H1, associé à une baisse de la teneur en antigène par unité de poids frais. Ces observations suggèrent qu’un enrichissement en CO2 atmosphérique et une forte intensité lumineuse sur le couvert végétal, bien que
favorables à la production de biomasse foliaire, ont un impact très limité sur le rendement net en antigène H1 au terme de la période d’expression hétérologue.
25
2.2 Abstract
Little is known about the influence of basic cultural practices on recombinant protein yields in plants used as bio-factories for biopharmaceuticals. We here assessed the effects of
atmospheric CO2 concentration and photosynthetic photon flux density (PPFD) on growth
and development of Nicotiana benthamiana used as a transient expression host for the
vaccine antigen influenza virus hemagglutinin 1 (H1). Our results showed CO2 enrichment
and high PPFD to increase leaf biomass and specific leaf weight in growth chamber conditions during the growth period before heterologous protein expression, but high PPFD decreased antigen content compared to total soluble protein on a leaf fresh weight basis. Expressed on a whole plant basis, soluble protein content was increased in plants grown under high PPFD but the H1 antigen yield following heterologous expression was only marginally influenced due to a negative impact on antigen content per leaf fresh weight. These observations suggest that CO2 enrichment and high PPFD, although promoting the
production of leaf biomass during plant growth, may have under some conditions a negligible impact on H1 yield per plant at the end of the protein expression period.
26
2.3 Introduction
Plants are attractive expression hosts for the heterologous expression of clinically- and industrially-useful proteins (Stöger et al., 2014; Tschofen et al, 2016). Plant-based systems offer several advantages for recombinant protein expression, such as short production cycles, low upstream production costs, convenient large-scale cultivation, the possibility to produce structurally-complex proteins, and low risks of pathogen or toxin contaminations as observed with animal and microbial cell expression hosts (Rybicki, 2010; Xu et al., 2012). Some plant-made proteins of medical interest are now on the market and several others are currently undergoing Phase II or Phase III clinical trials (De Muynck et al., 2010; Yusibov et al., 2011). A range of plants are used as expression hosts, notably including the wild relative of tobacco,
Nicotiana benthamiana (Lomonossoff and D’Aoust, 2016). This plant from Australia is a
convenient protein expression host in academic and industrial expression setups given its rapid growth rate, its large leaf biomass per plant and its natural ability to express foreign genes (Goodin et al., 2008; Wang and Bennetzen, 2015). Efficient transient expression systems have been developed in recent years for recombinant protein expression in N.
benthamiana, often involving vacuum infiltration of leaf tissue with the bacterial transgene
vector Agrobacterium tumefaciens (Leuzinger et al., 2013).
From a production standpoint, the yield of a plant-made biopharmaceutical depends on two parameters: (1) the amount of recombinant protein per biomass unit (in mg [or units]·g fresh weight–1); and (2) the biomass per plant or per growing area (in g·plant–1, or g·m–2). Many authors have proposed ways to improve recombinant protein production rates per plant biomass unit, often involving the development of molecular tools or strategies for increased transgene expression or protein stability (Fiedler et al., 1997; Cañizares et al., 2005; Robert et al., 2013; Hamorsky et al., 2015). By contrast, the influence of basic cultural conditions
27
on plant growth, biomass production and protein content have received little attention in a ‘molecular pharming’ context (Fujiuchi et al. 2014; Shang et al., 2018). Extensive work has been done over the years to determine optimal growth conditions for leafy crops like tobacco but the best cultural conditions for N. benthamiana expressing heterologous proteins may differ, as the ultimate goal is recombinant protein yield in leaf tissue, not total biomass production or increased endogenous protein content.
A widely used cultural practice known to promote growth and development of Solanaceae species and other C3 plants in controlled environments is CO2 enrichment, reported in many
instances to increase leaf biomass, relative growth, net carbon assimilation and specific leaf weight (e.g. Yelle et al., 1990; Ainsworth and Long, 2005; Kirschbaum, 2011). In theory, the time required for a C3 plant like N. benthamiana to reach a leaf biomass of 30 or 40 g suitable
for leaf agroinfiltration could be reduced upon CO2 enrichment, in such a way as to shorten
the plant’s upstream growth period before leaf infiltration for heterologous protein expression. Because high CO2-grown C3 plants tend to be sturdier, and their leaves to be
thicker, than plants cultivated under ambient CO2 concentration (Tremblay and Gosselin,
1998; Yin, 2002), a high-CO2 environment could also prove useful for N. benthamiana used
as a transient expression host by reducing leaf biomass losses inherent to plant handling and infiltration operations in vacuum chambers.
Supplemental lighting is another cultural practice likely having a positive impact on N.
benthamiana leaf biomass production, as light intensity directly determines the amount of
light intercepted and absorbed by the plant canopy. Supplemental lighting devices to implement high photosynthetic photon flux density (PPFD) regimes are particularly well suited to plant cultivation in controlled environment (e.g. in growth chambers), as they allow for a fine control of light amounts actually reaching the plant canopy. Supplemental lighting