Use of a continuous-flow bioreactor to evaluate nitrate reduction rate of Halomonas desiderata in cementitious environment relevant to nuclear waste deep repository
11
0
0
Texte intégral
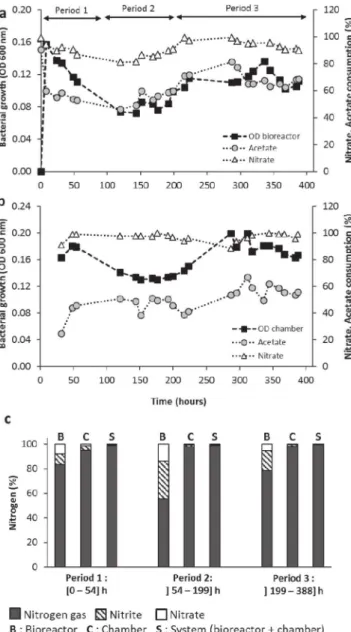
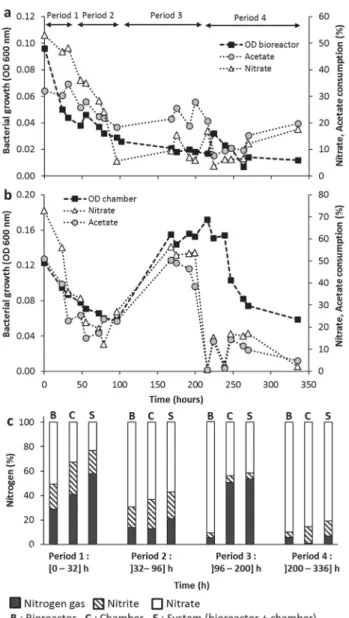
Figure

Documents relatifs
