Contrôle du développement embryonnaire par des petits ARN issus de transposons
Texte intégral
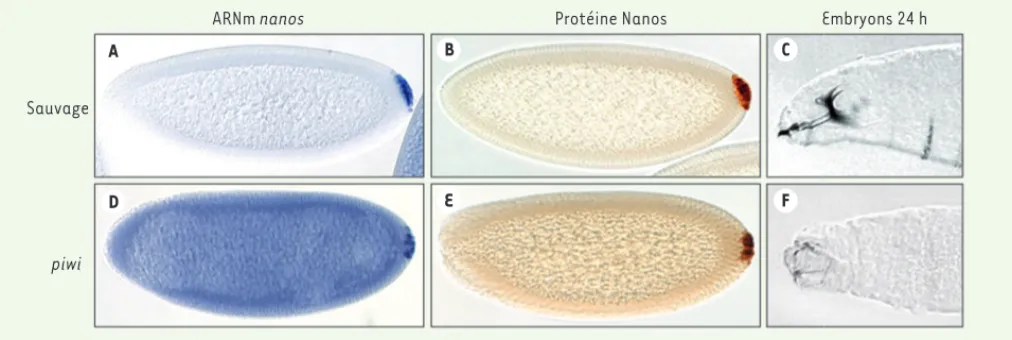
Figure

Documents relatifs
Cette quatrième série d’essais de co-expression révélait aussi la présence d’un complexe formé entre les protéines Snu13p et Pih1p (Figure 64, piste 129) et entre les
Comme mentionné dans les résultats supplémentaires, une des conclusions de notre travail est que le post conditionnement ischémique, efficace chez la souris à 2 mois, ne l’est
A comparison of the observed distribution of channel openings (Fig.3E) to that expected from the binomial distribution of six independently gating low N channels (Fig. 3C) shows
Pour un défaut de circuit-ouvert d'un transistor d’un onduleur, deux stratégies différentes sont proposées : la première est basée sur l’annulation des courants de
Si au contraire, c'est la régulation énergétique qui prédomine, la prise de glucides lipides et protéines va glisser le long de la pente iso- énergétique, la prise
Statistical analysis consisted of separate mixed-design analyses of variance (ANOVA) for the depen- dent variables (cumulative fixation duration and mean fixa- tion duration) and
1. Türbel besaß 1520 eine Kapelle mit Pfründe, die uun Stalden abhängig war. 1686 fand die endgültige Trennung statt. Petrus und Paulus).. Nach der Sage, die in Chroniken erwähnt
• Mais également c'est le lieu d'une passage massif des traditions écrites en chinois classique vers des textes en langue vulgaire. ➜ théâtre ➜ roman
