Contrôle β-adrénergique du courant calcique de type L cardiaque - Le chaînon manquant enfin découvert
Texte intégral
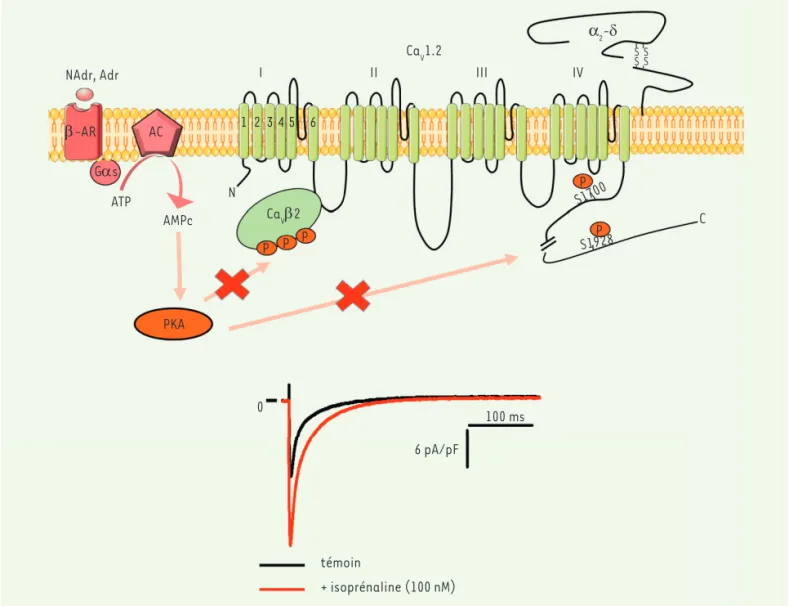
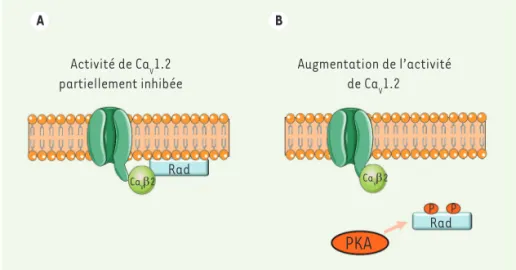
Figure
Documents relatifs
,CQ A?PBGMKWMAWRCQ QMLR JCQ ACJJSJCQ OSG BCK?LBCLR SL ?NNPMTGQGMLLCKCLR CL bLCPEGC GKNMPR?LR CR
1. Um welche Konzepte geht es in der Literaturüberprü- fung? Ganz besonders zu beachten sind die Konzepte der unabhängigen Variablen und abhängigen Variablen und ihre
Les réflexions très nombreuses sur le fonctionnement des marchés, en particulier le marché du travail mais aussi celui que constitue le partage de la production entre les
Pour un défaut de circuit-ouvert d'un transistor d’un onduleur, deux stratégies différentes sont proposées : la première est basée sur l’annulation des courants de
Tissier and coworkers published an extensive non-perturbative renormalisation group study of frustrated spin systems in dimension between two and three: a consequence of their study
D’une part, en s’inscrivant dans le mouvement d’effervescence qui avait gagné tous les intellectuels en Europe, le constitutionnaliste napolitain avait minutieusement examiné les
festonnés– dans– la– survenue– de–
Training the SVM models for group A, including calibration, on the 60,000 training sentences takes 7 minutes and 33 seconds for the best model (character 5grams), and 31 minutes
