The protein folding challenge in psychrophiles: facts and current issues
Texte intégral
Figure
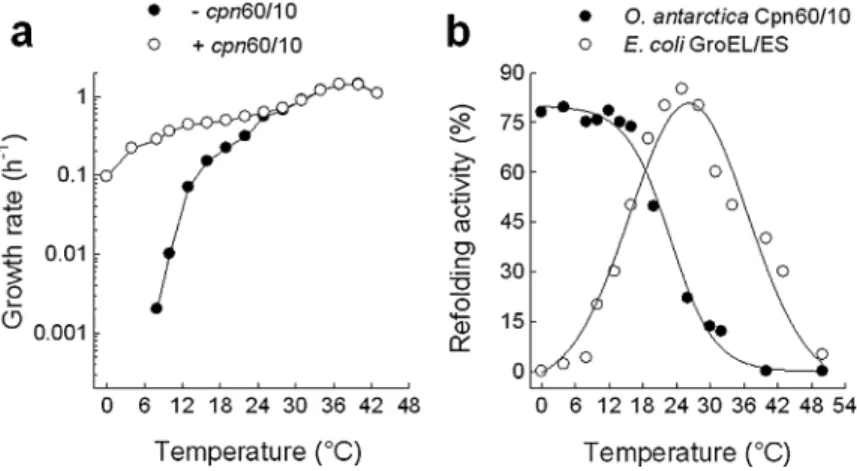
Documents relatifs
Interestingly, we find that the electrostatic analogy extends to defect chains: positive scars screen Gaussian curvature on the outer rim; negative scars appear in the inner region;
Methods Between 2005 and 2007 four patients (mean age 38.5 years, range 16–62 years) were treated with the device, in three patients to achieve parent artery occlusion of the
(2007) Tandem autologous stem cell transplanta- tion for patients with primary refractory or poor risk recurrent Hodgkin lymphoma. (2000) Prognostic factors and treatment outcome
Although the shape of a folded rod looks very complicated, elementary parts of the rod, delimited by two neighbouring junction points (where the rod is locally in self-contact) can
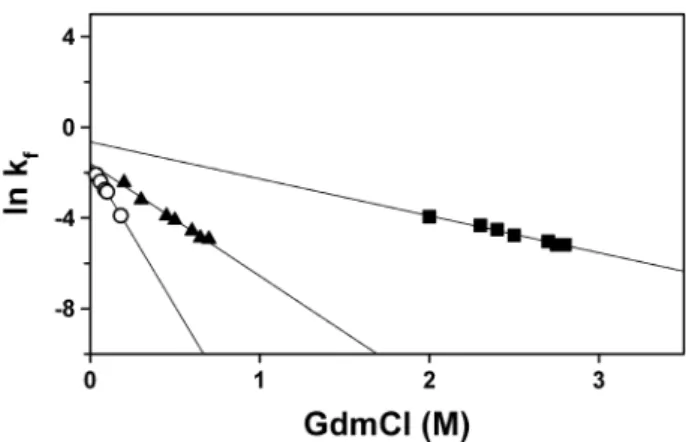
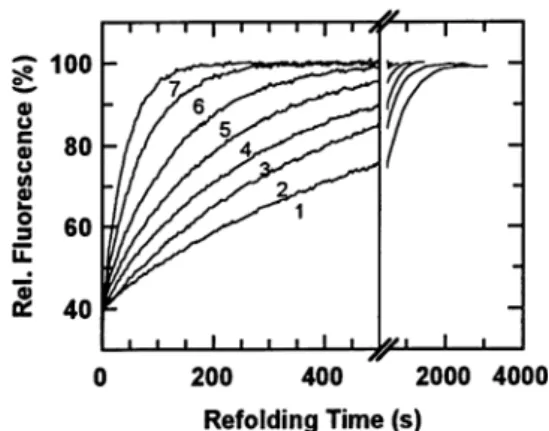
In another example, the acid-denatured states of ribonuclease, lysozyme and chymotrypsinogen, all three common model globular proteins for the study of protein folding, have shown
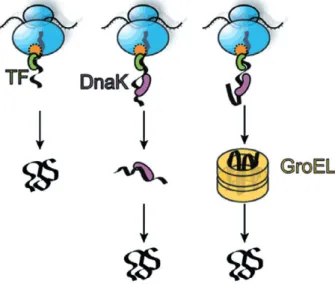
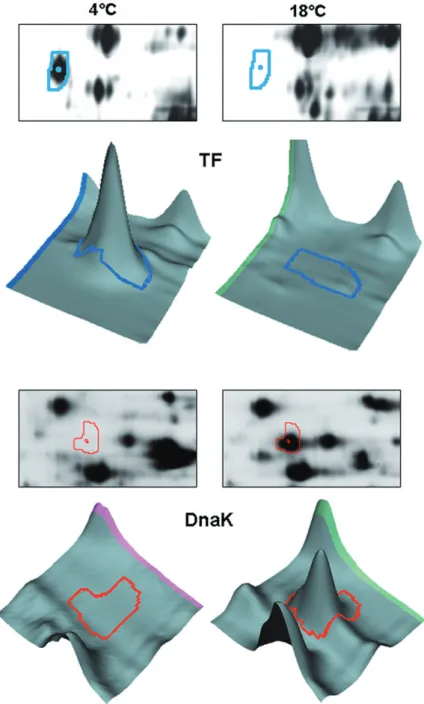
As previously mentioned, one of the main difficulties of the protein folding problem is the existence, in phase space, of a large number of local
For non chiral chains, a mean field study of this model shows that a classical 6 collapse transition occurs first; at lower temperature,.. nematic order
With a simple definition based on the well-known NKModel, the motivation is to make the IFP problem more accessible to optimisation specialists and model experts.Furthermore
