Patterns of diversification in early eukaryotes
Texte intégral
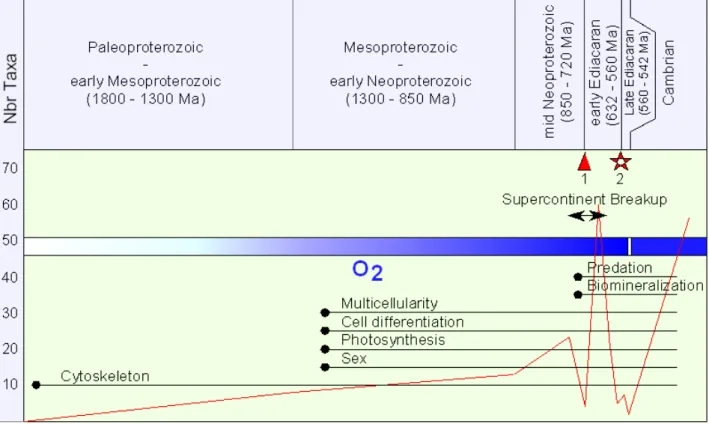
Figure
Documents relatifs
[r]
[r]
More precisely, the strict concavity of E v (N) will enable us to prove the nondegeneracy of the symplectic form restricted to the manifold of solitary waves.. Proof of Theorem 2.1.
Coatings of the comb polymer poly(L-lysine)-g- poly(ethylene glycol) (PLL-g-PEG) were investigated for potential to inhibit 1) nonspecific spreading of human blood-derived macro-
state that ‘scenario II’ (immediate surgery with sentinel lymph node biopsy for BI-RADS 5 lesions presenting as mass densities) could prevent 65% (i.e. 154 out of 238) of
Afterwards we examine in Section 4 the influence of the underlying lattice on the sublattice of representable weak complementations and then establish the characterization of
The SDRI-based tracking and positioning system we implemented on top of two core middleware services is fault-tolerant with respect to individual tag failures: (1) it redundantly
In summary, we have established an extraction se- quence of the group-4 elements from 8 M HCl into TBP, Zr > Rf > Hf, which is in qualitative agreement with
