De la communauté à l’individu: influence de l’intensité
agricole et du paysage sur les bourdons (Bombus spp.)
du Sud du Québec
Thèse
Amélie Gervais
Doctorat en biologie végétale
Philosophiæ doctor (Ph. D.)
De la communauté à l’individu : influence de
l’intensité agricole et du paysage sur les
bourdons (Bombus spp.) du Sud du Québec
Thèse
Amélie Gervais
Sous la direction de :
Valérie Fournier, directrice de recherche
Marc Bélisle, codirecteur
Résumé
Les pollinisateurs sont essentiels au maintien des écosystèmes naturels ainsi que pour la production d’une grande majorité de fruits et de légumes. Ils sont cependant globalement en déclin et de multiples facteurs sont mis en cause dont les changements climatiques, les maladies et parasites, la perte d’habitats naturels ainsi que l’augmentation des proportions de monocultures et de cultures intensives dans le paysage. Un gradient d’intensité agricole important parcourt le Sud du Québec d’ouest en est, d’agriculture très intensive à l’ouest vers un mode de production plus extensif à l’est. Dans ce contexte particulier, il devient intéressant de voir comment les bourdons, pollinisateurs essentiels de plusieurs cultures économiquement importantes à la région, réagissent et s’adaptent à un tel gradient. Pour tester l’importance des composantes du paysage sur les bourdons, plusieurs niveaux d’organisations ont été évalués, soit 1) les individus, 2) les colonies et 3) les communautés. Pour les niveaux des individus et des colonies, une expérience ayant eu lieu dans le Sud du Québec au printemps/été 2016 a été effectuée en installant 80 colonies commerciales de Bombus impatiens dans 20 paysages différents et étalés dans le gradient. Les paysages ont été caractérisés sur place, à un rayon de 1 km des colonies avec l’utilisation d’orthophotos et les catégories suivantes ont été formées (cultures intensives, cultures extensives, cultures à fleurs, forêts, milieu urbain et milieu aquatique). Hebdomadairement, l’activité des ouvrières à la colonie était évaluée et durant la saison, quatre ouvrières par colonie ont été capturées, mesurées et ont eu leur charge pollinique identifiée au niveau taxonomique le plus bas. Les résultats montrent que la taille des ouvrières n’est pas influencée par le contexte paysager. Leur quête alimentaire, quant à elle, semblait affectée, puisque les ouvrières présentes dans les paysages plus intensifs faisaient moins de va-et-vient à l’entrée de la colonie et ne transportaient pas les mêmes espèces de pollen que celles présentes dans les milieux plus extensifs. Les colonies, quant à elles, étaient suivies hebdomadairement et à chacune des visites, leur poids était mesuré. À la fin de leur vie, le poids final du nid, le nombre de reines produites ainsi que la présence d’Aphomia sociella étaient notés. Les colonies placées dans des paysages avec une plus grande proportion de cultures intensives ont vécu semaines de moins
et atteint un poids final et maximal moins important que les colonies placées dans des paysages plus extensifs. La proportion de cultures à fleurs a eu, quant à elle, un effet positif sur la longévité ainsi que sur les poids maximal et final des colonies. Le nombre de reines produites ainsi que la probabilité d’être déprédaté par A. sociella ne semblait pas être lié au paysage. Pour évaluer les communautés, 40 fermes situées dans le même gradient d’intensité agricole ont été échantillonnées de 2006 à 2016. Un total de 4726 reines bourdons, représenté par 13 espèces a été capturé à l’aide de pièges-fenêtre jaunes. Le paysage, décrit également par orthophotos, mais à 500 m de rayon cette fois, a influencé partiellement les communautés. Les proportions de cultures intensives, de cultures à fleurs ainsi que de forêts dans le paysage ont chacune influencé l’abondance et l’occurrence de quelques espèces, mais leurs effets n’étaient pas majeurs dans l’explication de l’assemblage des communautés. Ce sont les traits fonctionnels des espèces, particulièrement la taille des reines ainsi que leur site de niche qui influençait leur présence dans le paysage. Les résultats présentés dans cette thèse apportent encore plus d’évidences quant à l’impact du paysage, particulièrement les conséquences négatives des cultures intensives, sur des pollinisateurs essentiels, les bourdons.
Abstract
Pollinators are essential to natural ecosystems and crop production. However, they are in decline and many causes are possible: climate changes, diseases, parasites, loss of natural habitats, loss of floral resources, landscape homogenization and agricultural intensification. In southern Québec, a west-east gradient, from intensively managed landscapes in the west to a more extensive production in the east is present. In this context, it is quite interesting to see how pollinators, such as bumble bees, cope with such an important agricultural gradient. To test the importance of agricultural landscape composition on bumble bees, we investigated three different levels of organization: 1) individual, 2) colonies and 3) communities. To evaluate the impact of the landscape on the individuals and colonies levels, we placed 80 colonies of Bombus impatiens on 20 different sites in the gradient during the spring and summer of 2016. Landscapes were characterized within 1 km from the colonies using orthophotos and the landscape components were categorized following: intensive crops, extensive crops, flowering crops, forests, urban areas and aquatic environment (lakes, rivers). Each week, we counted the number of workers entering and exiting colonies and a total of four workers per colony were captured, measured and had their pollen load identified. Worker size was not influenced by the landscape. However, their foraging behavior seemed affected since the ins and outs of workers dropped in more intensive landscape. Furthermore, pollen species collected by workers were different in more intensive landscapes compared to those of more extensive landscapes. For the colony level, each one had its weight taken every week and at its death, its final weight, the number of queens produced and the presence of Aphomia sociella were noted. Colonies placed in the landscape with a high proportion of intensive crops gained less weight and lived almost two weeks less than colonies in more extensive landscape. The proportion of flowering crops in the landscape had the opposite effect by increasing both the weight and the life expectancy of colonies. The number of queens produced and the probability of being predated by A. sociella were not linked to landscape composition. At the community level, another study took place from 2006 to 2016 on 40 farms from the same agricultural gradient. A total of 4726 bumble bee queens were captured using yellow
window traps. The landscape was described again using orthophotos and the same categories, but at a 500 m radius instead. The proportions of intensive and flowering crops as well as the proportions of forests each influenced the abundance and occurrence of some species, but they were not the most important variables explaining the bumble bee community structure. The functional traits were, in fact, more important to explain the abundance of species, where smaller and underground nesters species were more abundant, in all the landscapes. The results presented in this thesis bring even more evidence that landscape has an important impact on bumble bees at every level. Furthermore, we highlight once more that intensive cropping systems are truly detrimental to bumble bee persistence in agroecosystems.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... vi
Liste des figures ... x
Liste des tableaux ... xiii
Remerciements ... xvii
Avant-propos ... xx
Introduction... 1
Chapitre 1 : États des connaissances ... 2
1.1. Les pollinisateurs indigènes ... 3
1.2. Le genre Bombus spp. ... 4
1.2.1. Ouvrières ... 5
1.2.2. Reines et colonies ... 7
1.2.3. Communautés ... 8
1.3. Déclin des pollinisateurs et principaux facteurs ... 9
1.3.1. Intensification agricole ... 10
1.3.2. Autres facteurs dans le déclin des bourdons ... 16
1.4. Le Sud du Québec ... 20
1.5. Problématique ... 21
1.6. Volet 1 —Les individus ... 22
1.7. Volet 2 —Les colonies ... 23
1.8. Volet 3—Les communautés ... 25
Chapitre 2 : Landscape composition and local floral resources influence foraging behavior, but not the size of Bombus impatiens Cresson (Hymenoptera: Apidae) workers ... 26 2.1 Résumé ... 27 2.2 Abstract ... 28 2.3. Introduction ... 29 2.4. Methodology... 33 2.4.1. Study sites ... 33
2.4.2. Colonies ... 33
2.4.3. Workers ... 34
2.4.4. Pollen loads ... 35
2.4.5. Landscape characterization ... 35
2.4.6. Local floral resources ... 36
2.4.7. Statistical analyses ... 37 2.5. Results ... 39 2.5.1. Morphology ... 39 2.5.2. Foraging Activity ... 40 2.5.3. Pollen ... 40 2.6. Discussion ... 41 2.6.1. Morphology ... 41 2.6.2. Foraging Activity ... 42 2.6.3. Pollen ... 44 2.7. Acknowledgement ... 46 2.8. References ... 47
Chapitre 3 : Agricultural landscape composition affects the development and life expectancy of colonies of Bombus impatiens Cresson (Hymenoptera: Apidae).... 64
3.1 Résumé ... 65 3.2 Abstract ... 66 3.3. Introduction ... 67 3.4. Methodology... 71 3.4.1. Model species ... 71 3.4.2. Study area ... 72 3.4.3. Quad monitoring ... 72
3.4.4. Worker pollen load ... 73
3.4.5. Landscape characterization and local floral resources ... 73
3.4.6. Statistical analyses ... 74
3.5. Results ... 78
3.5.1. Quad weight ... 78
3.5.3. Queen production ... 79
3.5.4. Nest weight ... 79
3.5.5. Depredation probability ... 79
3.5.6. Pollen load diversity and composition ... 80
3.6. Discussion ... 80
3.6.1. Landscape habitat composition ... 81
3.6.2. Local floral resources ... 84
3.7. Conclusion ... 85
3.8. Acknowledgement ... 86
3.9. References ... 86
Chapitre 4 : Impact of agricultural landscape composition on bumble bee communities in Southern Québec: a 10-year story ... 104
4.1 Résumé ... 105 4.2 Abstract ... 106 4.3. Introduction ... 107 4.4. Methodology... 111 4.4.1. Study area ... 111 4.4.2. Bumble bees ... 112 4.4.3. Landscape Composition ... 112 4.5. Results ... 114 4.6. Discussion ... 116 4.7. Acknowledgement ... 120 4.8. References ... 120 Conclusion... 136
Synthèses des résultats ... 137
Effet de la proportion de cultures intensives dans le paysage ... 137
Effet de la proportion de cultures à fleurs dans le paysage ... 140
Effet de la proportion de forêts dans le paysage ... 141
Limites de l’étude ... 143
Apport du doctorat à la communauté scientifique ... 146
Bibliographie ... 151
Annexe 1 ... 168
Annexe 2 ... 174
Liste des figures
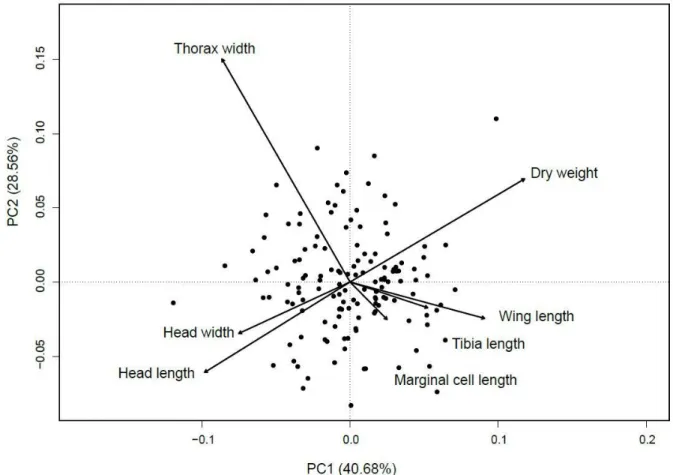
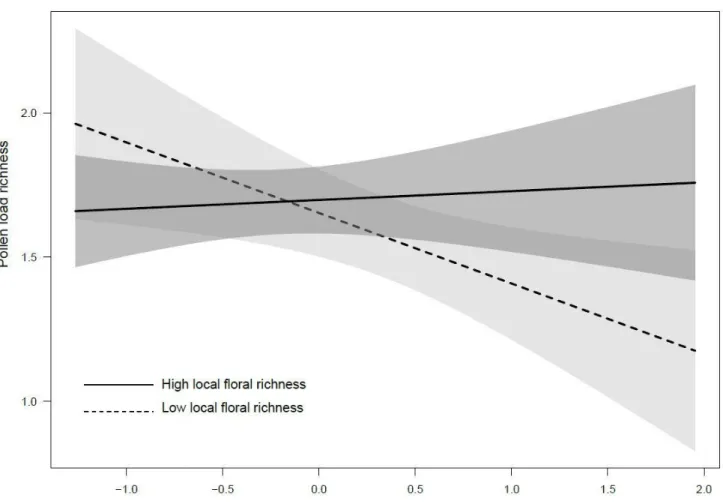
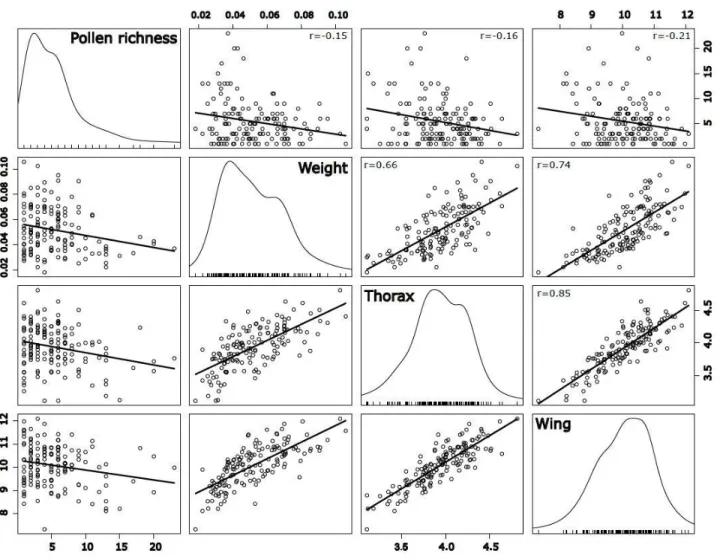
Figure 1. Habitat composition within 500 m of the 20 clusters (quads) of experimental Bombus impatiens colonies monitored in 2016 in Southern Québec, Canada ... 58 Figure 2. Principal Component Analysis (PCA) plot showing the multivariate variation among 205 bumble bee workers in terms of their morphometric measurements. Vectors indicate the direction of the functional trait to the overall distribution. The first two principal axes explained 69.24% of the variance. ... 59 Figure 3. Influence of intensive crop covers (500 m radius) on the foraging activity (entries and exits in 15 min) by the 20 clusters composed of 4 experimental colonies (quads) of Bombus impatiens monitored in 2016 in Southern Québec, Canada. Model-averaged predictions under average conditions are shown with 95% unconditional confidence intervals. ... 60 Figure 4. Species richness found in pollen load of 205 Bombus impatiens workers captured during the experiment in 2006 in function of the interaction between the proportion of flowering crops and the floral local diversity ... 61 Figure 5. Difference (based on Hellinger distance) in species between pollen load of workers and habitat in function of the interaction between the proportion of intensive crops and flowering crops. Workers were captured from 20 clusters composed of 4 experimental colonies of B. impatiens monitored in Southern Québec, Canada in 2016.. ... 62 Figure 6. Matrix of correlation between pollen richness, weight, thorax width and wing length of 205 workers of Bombus impatiens captured from 80 colonies placed on 20 sites during the summer of 2016 in Québec, Canada. ... 63 Figure 7.Habitat composition within 500 m of the 20 clusters (quads) of experimental Bombus impatiens colonies monitored in 2016 in Southern Québec, Canada. ... 97 Figure 8. Influence of intensive crop cover (500 m radius) on the survival rate of 68 experimental Bombus impatiens colonies clustered into 17 agricultural sites monitored in 2016 in Southern Québec, Canada. ... 98 Figure 9. Influence of intensive (a) and flowering (b) crop covers (500 m radius) on the temporal weight profiles of 17 clusters composed of 4 experimental colonies (quads) of Bombus impatiens monitored in 2016 in Southern Québec, Canada ... 99
Figure 10. Influence of intensive (a) and flowering (b) crop covers (500 m radius) on the maximum weight attained by the 17 clusters composed of 4 experimental colonies (quads) of Bombus impatiens monitored in 2016 in Southern Québec, Canada ... 100 Figure 11. Forest plot of model-averaged coefficients and their 95% unconditional confidence intervals estimated by multimodel inference following model selection (see Table 5). Model coefficients were computed with standardized explanatory variables (zero mean and unit variance) ... 101 Figure 12. Influence of flowering crop cover (500 m radius) (a) and local flower species richness 100 m radius) (b) on the nest weight at death of 68 experimental Bombus impatiens colonies clustered into 17 agricultural sites monitored in 2016 in Southern Québec, Canada.. ... 102 Figure 13. Influence of landscape habitat composition and local floral resources on the species composition of pollen loads collected by the workers of 68 experimental Bombus impatiens colonies clustered into 17 agricultural sites monitored in 2016 in Southern Québec, Canada. ... 103 Figure 14. Habitat composition of the 40 monitored from 2006 to 2016 in Southern Québec, Canada. ... 128 Figure 15. Landscape components for the 11 years of the study. a) Proportion of intensive crops; b) Proportion of flowering crops; c) Proportion of forests. Each point represents a farm at a given year. ... 129 Figure 16. Bayesian joint model of the global community ... 130 Figure 17. Phylogenetic signal computed from the two bayesian joint models: The global community and the queen only. ... 131 Figure 18. Estimates and confidences intervals computed from both Bayesian joint models illustrating the size effect of the proportion of intensive crops, flowering crops and forests on bumble bee functional traits.. ... 132 Figure 19. Bayesian joint model of the queens only.. ... 133 Figure 20. Number of queens captured during the course of the 11 years of the study
Figure 21. Number of queens captured during the course of the study as a function of their nesting preference. ... 135 Figure 22. Schéma illustrant différentes variables et relations importantes pour l’écologie du bourdon ... 143 Figure 23. Correlation matrix of the landscape composition and configurations variables ... 174
Liste des tableaux
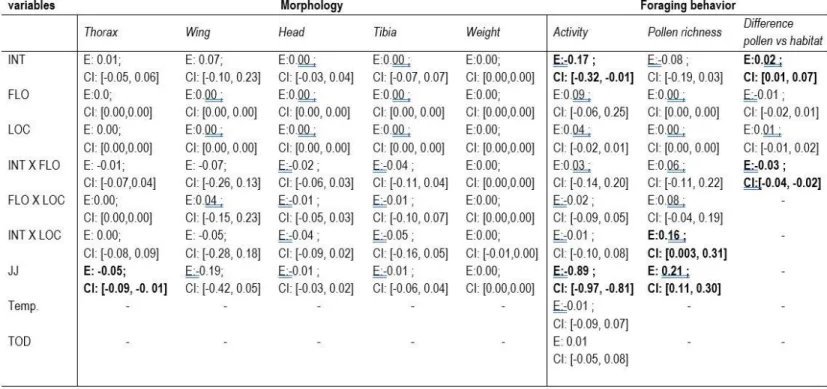
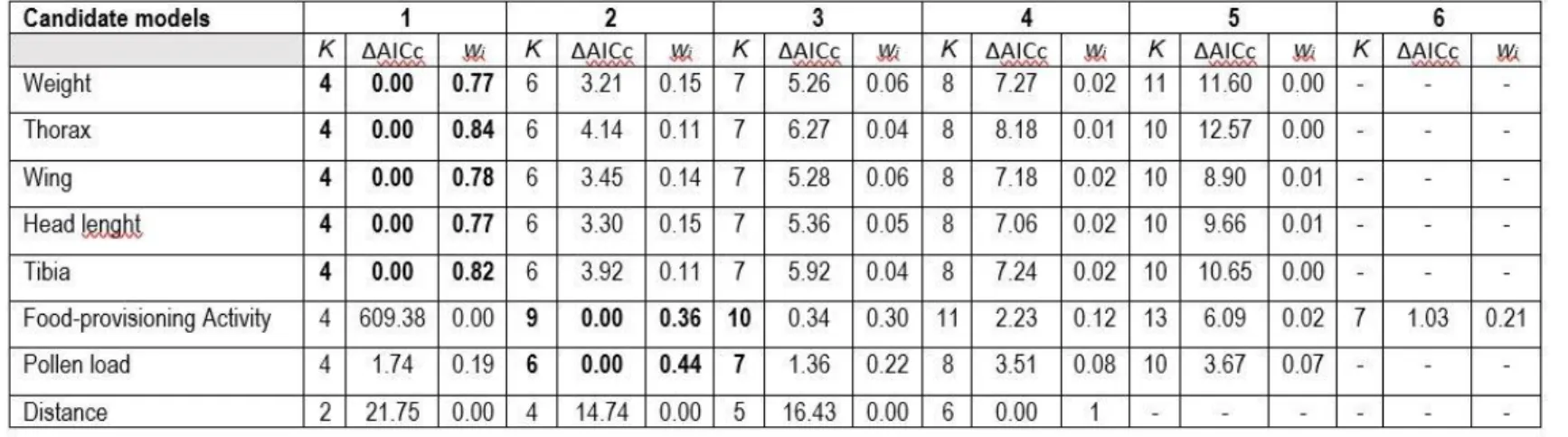
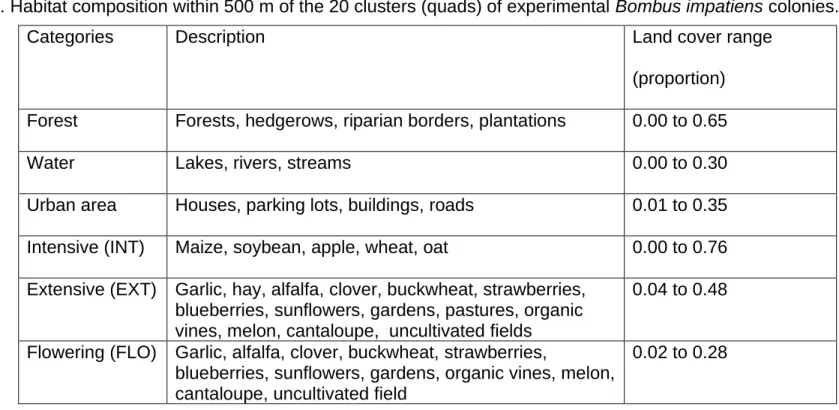
Table 1. List of all the candidate models used to model-average the estimates. INT: Proportion of intensive crops; FLO: Proportion of flowering crops; LOC: Local floral richness. ... 55 Table 2. Model-averaged coefficients and their unconditional 95% confidence intervals estimated by multimodel inference following model selection (see Table 1). Model coefficients were computed with standardized explanatory variables (zero mean and unit variance). ... 56 Table 3. Candidate models considered in model selection and multimodel inference procedures based on the second-order Akaike information criterion (AICc). K: number of model parameters; wi: Akaike weight; Candidate models (from 1 to 6) are presented in Table 1. ... 57 Table 4. Habitat composition within 500 m of the 20 clusters (quads) of experimental Bombus impatiens colonies. ... 93 Table 5. List of all the candidate models used to model-average the estimates.... 94 Table 6. Outputs of the two generalized additive mixed models (fitted with mgcv in R) used to quantify the (univariate) influence of intensive and flowering crop covers (500-m radius) on the temporal weight profiles of 17 clusters composed of 4 experimental colonies (quads) of Bombus impatiens monitored in 2016 in Southern Québec, Canada. ... 95 Table 7. Candidate models considered in model selection and multimodel inference procedures based on the second-order Akaike information criterion (AICc). K: number of model parameters; wi: Akaike weight; INT: Proportion of intensive crops;
FLO: Proportion of flowering crops; EXT: Proportion of extensive crops; LOC: Local floral resources (species richness). ... 96 Table 8. Parameters estimates and 95% confidence interval (LB= Lower bound; UB= Upper bound) of the model looking at the abundance of the queens captured in Quebec, Canada, for the 11 years of the project in function of their species functional traits. ... 127 Table 9. ∆AICc comparing both scale (500m vs 1000m) for all models. ... 168
Table 10. List of the plants found in the pollen loads of workers. # workers represent the number of workers carrying this pollen (n= 264) ... 169
«The greater our knowledge increases, the greater our ignorance unfolds.» - John F. Kennedy
Remerciements
Quand on entreprend un projet de l’ampleur d’un doctorat, il y a certainement plusieurs personnes qui ont contribué directement et indirectement à sa réussite. J’aimerais d’abord remercier ma directrice, Valérie Fournier, qui m’a accompagnée pendant presque sept ans durant mon passage aux études graduées. Merci de m’avoir guidée et de m’avoir donné toutes les opportunités possibles pour m’épanouir dans ma future carrière. J’aimerais aussi remercier mon co-directeur, Marc Bélisle, de m’avoir aidée à développer mon esprit critique, mon sens de l’organisation (mes parents ont essayé pendant de nombreuses années et n’ont pas réussi), particulièrement dans mes scripts R et de m’avoir aidé à devenir, je l’espère, une meilleure scientifique.
J’aimerais remercier mes parents de m’avoir partagé leurs amours, leurs valeurs, de m’avoir stimulée, soutenue et d’avoir répondu à mes milles-et-une questions quand j’étais enfant. C’est grâce à vous que j’ai cette grande curiosité qui me permet d’être créative et qui me fait autant aimer la recherche. Vous avez encouragé mes passions (sauf l’archéologie.), c’est le plus beau cadeau que vous puissiez me faire. Ce doctorat est aussi à vous, merci. Merci aussi à mon bébé sœur, la plus belle des Isabelles, la meilleure des physios, la meilleure des petites sœurs. Merci d’avoir été là pour moi, de toujours m’avoir poussée à faire mieux, et ce, dans tous les domaines inimaginables (ne pas oublier mon lunch ou même me faire un bon lunch, faire mes « cassettes», écrire de bonnes lettres de motivation en me vendant mieux, etc..). Je n’ai aucune idée comment je faisais quand tu n’étais pas là, une chance que je t’aie. Un gros merci aussi à ma famille plus élargie d’avoir été présente pour moi. On ne se voit pas souvent, juste pour des occasions joyeuses, ou moins joyeuses, mais je me sens toujours aimée et soutenue en votre présence. Ce sentiment a certainement contribué à la réussite de ce doctorat. Merci mononcle C’Alain, mononcle Jacques, matante Lisette, matante Hélène, matante Lise, matante Nicole et mononcle André, les cousins et les cousines, je vous aime !
J’aimerais rajouter un petit mot pour ma grand-mère maternelle qui n’a malheureusement pas pu voir l’aboutissement de ce beau projet. Chaque année, elle donnait à tous ses petits et arrière-petits-enfants 100 $ pour Noël s’ils allaient à l’école. C’était toute qu’une avant-gardiste ! C’est sans aucun doute moi qui lui ai coûté le plus cher dans la famille, mais elle n’a jamais arrêté de m’encourager. Merci, grand-mère, je t’aime.
J’aimerais remercier une personne bien spéciale pour moi qui m’a soutenue tout au long de la maitrise et du doctorat. Il a partagé ma vie pendant presque 9 années. Nos chemins se sont séparés avant la fin de ce projet, mais sans lui, ces quatre dernières années auraient été bien pénibles. Merci, Louis-Philippe, d’avoir été mon partenaire pour les bons et moins bons moments pendant toutes ces années.
Merci à mes deux grands frères adoptifs, qui m’ont aidé et encouragé tout au long de mes études graduées. J’ai beaucoup apprécié vos conseils et j’adore nos discussions. Je me trouve vraiment choyée d’avoir pu suivre vos traces dans le fantastique monde des pollinisateurs, merci Joseph et Étienne.
I would like to thank you, Cory, from the bottom of my heart. Thank you for your never-failing support, your advice and your presence in my life. My master’s would have been painful to do, and I would not have had the necessary tools to finish this PhD if it wasn’t for you. Thank you.
J’aimerais terminer en soulignant à quel point je suis chanceuse d’avoir des amis incroyables qui m’ont permis de garder une santé mentale saine (enfin je crois) et les pieds sur terre pendant ces quatre années. Que ce soit par des sorties, du sport ou des soirées d’études, votre présence m’a grandement aidée. Parmi ceux-ci, ma « gang » de volley qui m’a tenu en forme les lundis soirs, merci, Pierre-Luc, Andréanne, Jean-Pierre, Julien et Yan, vous êtes « amazing ». Merci à ma « gang » de lab d’avoir su décrypter tous mes trucs pour éviter de sortir et d’avoir trouvé une façon d’y contrecarrer par diverses méthodes comme la culpabilité, le « peer
pressure » et le « ça parait bien dans un CV ». Mon CV et mes souvenirs seraient certainement moins remplis si vous n’aviez pas utilisé vos fameuses tactiques, merci à Stéphanie, Marianne, Sabrina, Mouna, Frédéric, Mélanie, Phanie, Olivier, Sandrine et Anne de votre soutien tout au long de ces années. Merci à la « gang du BAC » de Sherbrooke expatriée à Québec qui a continué de m’inviter à des activités malgré mes innombrables refus. Votre présence, votre support et votre énergie ont beaucoup compté pour moi, plus que vous ne le pensez, merci, Gaboche, Charlou, Andréane et Mathieu. J’aimerais aussi remercier mes trois mousquetaires qui ont été là pour moi durant des moments moins faciles, qui m’ont fait suer en masse au squash et qui m’ont fait boire du vin au prix du cost, je vous adore, merci d’être là Olivier, Simon et Benoit.
Avant-propos
Cette thèse est composée de cinq chapitres, incluant une introduction et une conclusion générales rédigées en français, ainsi que trois chapitres écrits en anglais sous forme d’articles scientifiques. Tout le contenu de la thèse présentée ici a été entièrement écrit par la candidate au doctorat, avec les commentaires et conseils de Valérie Fournier (Université Laval, Département de phytologie) et Marc Bélisle (Université de Sherbrooke, Département de biologie), directrice et co-directeur.
Le premier chapitre est une introduction générale présentant les principaux enjeux concernant les pollinisateurs indigènes, et traitant plus particulièrement des bourdons dans le paysage agricole. C’est aussi dans ce chapitre que les hypothèses et objectifs de recherche sont présentés.
Le chapitre 2, soit le premier article de recherche de la thèse, s’intéresse à l’effet de la composition du paysage sur la morphologie et la quête alimentaire d’ouvrières de Bombus impatiens Cresson. Ce chapitre intitulé « Landscape composition and local floral resources influence foraging behavior, but not the size of Bombus impatiens Cresson (Hymenoptera: Apidae) workers » a été soumis pour publication à la revue Plos One en octobre 2019. La préparation de l’expérience, les analyses statistiques, ainsi que la rédaction de l’article, ont été effectuées par la candidate, avec le soutien de sa directrice et co-directeur. Ève Courtois, co-auteure, a participé à une partie importante de la prise de données et a soumis ses commentaires pour la rédaction de l’article.
Le chapitre 3 est un article de recherche qui s’intéresse également à l’effet du paysage agricole, mais sur un niveau d’organisation plus élevé, soit les colonies de B. impatiens. Ce chapitre intitulé « Agricultural landscape composition affects the development and life expectancy of colonies of Bombus impatiens Cresson » a été soumis pour publication à la revue Ecosphere le 5 août 2019 et accepté le 18 mars 2020. Encore une fois, la planification de l’expérience, la collecte de données,
l’analyse des résultats et la rédaction de l’article ont été principalement faites par la candidate, avec les commentaires et conseils de Valérie Fournier et Marc Bélisle.
Le chapitre 4 et dernier article de recherche de la thèse visait à quantifier l’effet de la composition du paysage sur l’abondance, l’occurrence et les traits fonctionnels des communautés de bourdons retrouvés dans le Sud du Québec. Il s’agit d’une étude qui s’échelonne sur plus de dix ans. Ce chapitre s’intitule « Impact of agricultural landscape composition on bumble bee communities in Southern Québec: a 10-year story ». Il est prévu de le soumettre pour publication en avril 2020 au journal Ecology. La prise de données, les analyses statistiques et la rédaction de l’article ont été principalement faites par la candidate. Marc Bélisle a participé à la planification de l’étude en 2006 et il a aussi partagé de nombreux conseils et commentaires lors de la rédaction de l’article. Valérie Fournier a également fourni ses commentaires pour la rédaction de l’article. Guillaume Blanchet a prêté son expertise statistique pour valider les résultats ainsi que la partie écrite des statistiques présentées dans ce chapitre.
Finalement, le chapitre 5 constitue la conclusion générale de la thèse. Il comprend une brève synthèse des résultats et une interprétation de l’impact du paysage sur les trois niveaux d’organisations évalués ici, soit les individus, les colonies et les communautés. Il présente également les limitations de la thèse, son apport à la communauté scientifique et les futures perspectives de recherche dans le domaine.
Introduction
Dans un environnement constamment en changement, les organismes qui y habitent doivent présenter une forte plasticité phénotypique ou s’adapter pour survivre (Chown et al., 2007). Cette réalité touche même les insectes qui sont bénéfiques aux humains, comme les pollinisateurs. Ces derniers sont reconnus pour participer à la pollinisation du tiers de la production mondiale en fruits et légumes (Klein et al., 2007). Cependant, les pressions combinées de plusieurs facteurs comme l’intensification agricole ou la perte d’habitats naturels menacent grandement ce service offert par les insectes (Rader et al., 2014 ; Smith et al., 2013). Il devient alors important de comprendre comment ces différents facteurs influencent les pollinisateurs indigènes pour pouvoir prendre des décisions plus éclairées pour leur conservation. Le Sud du Québec, qui inclut la Montérégie et l’Estrie, représente le berceau de l’agriculture de cette province. En effet, cette région possède un gradient agricole très important, passant de l’agriculture très intensive à l’ouest, vers une agriculture plus extensive à l’est (Jobin et al., 2003 ; Bélanger and Grenier 2002). Cette variation d’intensité retrouvée au niveau du paysage influence certainement les différentes communautés de pollinisateurs indigènes.
L’objectif principal du projet est de quantifier comment la composition du paysage, comme proxy de l’intensité agricole, affecte l’écologie des bourdons (Bombus spp.). Le premier volet s’attarde à un petit niveau d’organisation, c’est-à-dire au bourdon lui-même. Le but de ce volet est de déterminer comment l’intensité agricole influence la taille et la quête alimentaire des individus. Le deuxième volet touche un plus grand niveau d’organisation, soit la colonie. Le but ici est de déterminer comment l’intensité agricole influence le développement et la reproduction de la colonie. Finalement, le troisième volet du projet vise à étudier l’écologie des communautés des bourdons. Le but est de déterminer si les assemblages d’espèces ou les traits fonctionnels des bourdons varient en fonction de la composition du paysage. L’observation simultanée de l’impact de ces facteurs sur plusieurs niveaux d’organisations est une approche novatrice pour étudier l’écologie du bourdon.
Chapitre 1 : États des connaissances
1.1. Les pollinisateurs indigènes
La pollinisation constitue un service écosystémique important, c.-à-d. un service offert par les écosystèmes aux humains. La pollinisation peut être abiotique, soit par le vent ou l’eau, ou elle peut être biotique, soit par les mammifères, les oiseaux et les insectes. De fait, près d’un tiers de la nourriture mondiale, soit 87 cultures, nécessite une pollinisation animale pour une production efficace de fruits, de noix, de grains ou de légumes (Klein et al. 2007). Ce service offert mondialement par les pollinisateurs était évalué à 204 milliards $CAN en 2009 (Gallai et al. 2009). Essentielle pour la reproduction sexuée des plantes, la pollinisation représente en fait une relation mutualiste entre la plante et ses pollinisateurs. Les pollinisateurs se nourrissent de la plante en récupérant pollen et nectar, des substances essentielles à leur survie et à celle de leur progéniture. Le pollen comble en effet leurs besoins en acides aminés, lipides et minéraux, alors que le nectar comble leurs besoins en glucides (de Groot 1952). Par le biais de leur quête alimentaire, les pollinisateurs amassent des grains pollen d’une plante ou fleur mâle et certains grains seront ainsi déposés, par accident, sur le stigmate d’une plante ou fleur femelle, assurant ainsi la reproduction sexuée de ces dernières.
Groupe formé de plusieurs ordres et familles taxonomiques, les pollinisateurs indigènes varient selon les systèmes étudiés. En Amérique du Nord, les insectes pollinisateurs font surtout partie des ordres des coléoptères, des lépidoptères, des diptères et des hyménoptères. Ces deux derniers ordres constituent les deux plus importants groupes de pollinisateurs à nos latitudes (Ssymank et al. 2008). Les diptères sont très diversifiés et leurs histoires de vie varient grandement entre les familles, les genres et même les espèces. Les Syrphidae et les Bombyliidae représentent les deux principales familles de diptères qui agissent comme pollinisateurs. Ils sont très efficaces là où les autres pollinisateurs échouent, c’est-à-dire en hautes latitudes (≥60°N ou S) et en hautes altitudes (Ssymank et al. 2008). Cependant, les pollinisateurs les plus importants font certainement partie de l’ordre des hyménoptères, plus particulièrement de la superfamille des Apoidea, soit les abeilles.
Le représentant des Apoidea le plus connu est sans aucun doute l’abeille à miel, aussi appelée abeille domestique (Apis mellifera L.). Cette dernière est bien connue du grand public pour sa production de miel et son importance dans la pollinisation des fruits, comme les pommes, les bleuets et les canneberges. Sa contribution économique annuelle à l’agriculture par le biais de ses services de pollinisation était estimée entre 3 à 5 milliards $CAN en 2013 (Darrach and Page 2016). Elle ne représente cependant qu’une seule espèce parmi les 730 espèces d’abeilles identifiées au Canada (Packer 2007). Il existe six familles d’Apoidea au Canada : Andrenidae, Apidae, Colletidae, Halictidae, Megachilidae et Melittidae (Packer 2007). Chacune de ces familles possède des caractéristiques qui lui sont bien particulières. La famille des Apidae, par exemple, est la seule famille qui inclut des espèces possédant des corbeilles à pollen. Cette corbeille, dont l’abeille à miel et le bourdon ont hérité, est une structure morphologique spécialisée qui leur permet d’amasser une grande quantité de pollen sur leurs pattes arrières.
1.2. Le genre Bombus spp.
Les bourdons (Bombus spp.), de la famille des Apidae, constituent les pollinisateurs les plus efficaces pour plusieurs cultures (Goulson 2010). Il est l’une des rares abeilles à effectuer la sonification des fleurs. Ce phénomène consiste à faire vibrer les anthères des fleurs à haute fréquence via un vrombissement des ailes afin que le pollen s’éjecte beaucoup plus efficacement. Cette technique est particulièrement utile pour les fleurs possédant des anthères porifères (c.-à-d., avec une seule petite ouverture par laquelle peut s’échapper le pollen), comme la tomate, le bleuet et la canneberge. De plus, la grande taille des bourdons, comparativement aux autres membres de la même famille, leur permet de polliniser même lorsque les températures sont froides (autour de 0°C). Cette qualité est particulièrement utile lorsque les cultures fleurissent tôt au printemps. Cependant, la contribution des bourdons aux écosystèmes agricoles est difficilement évaluable et probablement sous-estimée. Le genre Bombus est relativement diversifié, avec 59 et 238 espèces en Amérique du Nord et dans le monde respectivement (Ascher and Pickering
2018). La biologie de chaque espèce diffère et très peu d’informations sont disponibles pour la plupart d’entre elles.
Deux espèces de bourdons ont été « domestiquées » pour les services de pollinisation, soit B. impatiens en Amérique du Nord et B. terrestris en Europe. Pour la tomate de serre en Europe seulement, la valeur du service de pollinisation par ces bourdons est estimée à 12 G€ par année (Velthuis and Van Doorn 2006). Cette domestication est rendue possible du fait que ces espèces de bourdons peuvent être élevées dans des conditions artificielles (boite en carton). Leur domestication a débuté dans les années 70, mais ce n’est qu’en 1987 que les colonies furent commercialisées (Velthuis and Van Doorn 2006). Contrairement à la majorité des autres espèces du même genre, la biologie de ces deux espèces est très bien comprise. Elles utilisent une grande variété de matériaux et de sites de nidification (p. ex. sous terre, dans les racines d’herbacées, dans les cavités, dans d’anciens nids de rongeur), chose qui facilite grandement leur commercialisation. Contrairement à l’abeille à miel, les colonies de bourdons ne produisent pas de miel et les ruchettes vendues servent exclusivement à la pollinisation. Les ruchettes « artificielles » de bourdons, souvent appelées « quad » lorsqu’elles sont en groupe de quatre, sont disponibles auprès d’entreprises spécialisées comme Koppert Biological System © et BioBest.
1.2.1. Ouvrières
Les bourdons sont considérés comme des insectes sociaux, c’est-à-dire qu’ils vivent en colonie et possèdent des castes différentes (reines, ouvrières et mâles). Les ouvrières, qui proviennent d’un œuf fécondé produit par la reine, participent à la majorité des tâches au sein de la colonie. Elles occupent les rôles de nourrices, ménagères et butineuses (Goulson 2010). Seule la reine est en charge de la ponte des œufs dans la colonie. Contrairement à l’abeille à miel qui voit sa répartition des tâches effectuée selon l’âge de l’ouvrière (ou polythéisme d’âge), la distribution des tâches chez les bourdons est moins connue. Les tâches pourraient être distribuées selon la taille des ouvrières (qui varient grandement à l’intérieur d’un même nid ;
Couvillon and Dornhaus 2009; Couvillon et al. 2010), où les plus petites ouvrières s’occuperaient des tâches reliées au nid, alors que les plus grosses ouvrières occuperaient le rôle de butineuses. Les petites ouvrières pourraient effectivement se déplacer plus efficacement à l’intérieur du nid, alors que les grosses ouvrières devraient, par leur taille, pouvoir voler plus loin, rapporter plus de ressources par voyage et voler par temps plus froid (Couvillon et al. 2010). Cette division des tâches appelée polythéisme de taille pourrait permettre aux bourdons de mieux s’adapter et/ou être le résultat d’un compromis entre des ouvrières de faible qualité peu « coûteuses » et des ouvrières « dispendieuses » de haute qualité (Couvillon et al., 2010 ; Goulson et al. 2002b ; Peat et al. 2005).
Les ouvrières butineuses ont l’importante tâche de ramener les ressources essentielles au bon développement de leur colonie : le pollen et le nectar. Les fleurs ne possèdent pas tous les mêmes éléments nutritifs, et pas toutes dans les mêmes proportions (de Groot 1952). De plus, la capacité de produire du nectar varie d’une espèce florale à l’autre. Cette variation dans la qualité nutritive des différentes ressources florales disponibles amène les butineuses à utiliser différentes stratégies de butinage. La colonie a avantage à multiplier les types de fleurs butinées par ses ouvrières, puisque chaque pollen offre des acides aminés essentiels différents (de Groot 1952; Somme et al. 2015). On devrait donc observer une diversification au niveau de la colonie, bien que ceci n’ait jamais été testé dans la littérature, à ma connaissance. En revanche, les ouvrières ont avantage, lorsque les ressources florales sont en assez grandes quantités, à concentrer leurs efforts sur le moins d’espèces florales possible (Raine and Chittka 2007). Les ouvrières, elles, devraient se spécialiser. Les fleurs possèdent toutes des formes différentes et des caractéristiques spécifiques, ainsi un temps d’apprentissage est nécessaire pour permettre aux ouvrières de retirer les ressources de manière efficace (Gegear and Laverty 2005; Grüter and Ratnieks 2011). Donc, dans un environnement où la nourriture est disponible en grande quantité, les ouvrières auront donc tendance à se spécialiser pour un nombre limité de fleurs. À l’inverse, si les fleurs sont
disponibles en quantité limitée, les ouvrières devront laisser tomber leur stratégie de spécialisation et devenir généralistes pour maximiser les ressources.
1.2.2. Reines et colonies
Les reines et les mâles sont les deux castes reproductrices de la colonie. C’est à la reine que revient la tâche d’assurer la pérennité de la colonie ainsi que sa succession en produisant ouvrières et futures reines (Goulson 2010). Elle y parvient en fécondant ses œufs par le sperme provenant de l’accouplement effectué avec un/des mâles d’autres colonies. Contrairement aux ouvrières et aux reines, les mâles sont le produit d’un œuf non fécondé (Goulson 2010).
Au Québec, il n’y a qu’une seule génération de bourdons par année. Les reines, produites à l’automne, émergent d’une diapause hivernale au printemps suivant. Elles se mettent alors à la recherche de nourriture et d’un nid qui pourra abriter leur future colonie. Le type de nid recherché par la reine, qui varie grandement entre les espèces, doit pouvoir contenir l’entière colonie pour la saison de croissance. Selon l’espèce, le nid peut être sous terre, sous une grosse pierre, dans une cavité dans un arbre, dans des nichoirs à oiseaux, etc. (Goulson 2010). De plus, la présence de nourriture à proximité de ce nid est primordiale à la survie de la colonie. Lorsque la reine est établie, elle amasse du pollen et du nectar pour sa consommation propre et celle de sa future progéniture. Lorsque suffisamment de pollen est amassé, la reine pond ses œufs, qui deviendront ultimement ses premières ouvrières. Elle continue à faire des va-et-vient pour chercher de la nourriture, jusqu’au développement complet de ses ouvrières (~24 jours) (Goulson 2010). Elle reste ensuite au nid à pondre, tandis que ses ouvrières fourniront soins et nourriture aux futures ouvrières pondues par la reine. La colonie prend alors du poids en amassant des réserves en quantité suffisante pour produire les futurs ouvrières, reines et mâles. Peu avant la fin de l’été, avant de mourir, la vieille reine commence à pondre les futures reines et mâles. Les mâles chercheront les nouvelles reines produites pour s’accoupler à la fin de l’été ou à l’automne, puis ces dernières chercheront un abri pour passer l’hiver (Goulson 2010). Les mâles meurent à la fin de l’automne.
1.2.3. Communautés
Plusieurs méthodes permettent de quantifier la diversité en espèces. Parmi les plus employées, les indices de Shannon et Simpson utilisent la proportion des espèces à chacun des sites pour évaluer respectivement l’homogénéité et la probabilité de rencontrer une nouvelle espèce dans la communauté d’intérêts (Magurran 2004). Ces indices ont l’avantage d’être simples, mais le désavantage d’engendrer une perte d’information importante en limitant la variation des espèces à une valeur unique. Les méthodes multivariées permettent, quant à elles, d’illustrer la variation retrouvée à chacun des sites (ex. analyse en composantes principales ; Legendre and Legendre 2012a) ou d’illustrer la variation entre les sites en relation avec des variables environnementales (ex. analyse de redondance canonique ; Legendre and Legendre 2012b). Ces types d’analyses permettent de condenser l’information sur chacune des espèces en rendant les résultats minimalement digestibles avec un minimum de pertes d’information.
Au Québec, on retrouve 27 espèces de bourdons (Williams et al. 2014). La plupart du temps, ces espèces cohabitent entre-elles en présentant des niches écologiques différentes (phénologie décalée, choix de fleurs et de site de nidification différents). La diversité de leurs différents traits fonctionnels, c.-à-d. traits morphologiques, phénologiques ou physiologiques qui leur donnent un avantage ou un désavantage selon le type d’environnement, permettent généralement de limiter la compétition interspécifique et d’assurer une communauté stable et diversifiée. Les traits morphologiques des bourdons peuvent être associés à des fonctions écologiques. Par exemple, la largeur du thorax et la longueur des ailes sont souvent associées à la distance de vol des bourdons (Walther-Hellwig and Frankl 2000; Westphal et al. 2006a; Kapustjanskij et al. 2007; Polidori et al. 2013). La longueur de la tête, liée à la longueur de la glosse (langue), permet d’identifier le type de fleurs préférablement visité par l’espèce (Harder 1982; Harder 1983; O’Neill and O’Neill 2011). La taille du tibia arrière, par exemple, pourrait quant à lui indiquer la taille de la corbeille à pollen et donc être liée à la quantité de nourriture ramenée à la colonie par voyage.
Ces traits morphologiques, combinés à d’autres traits écologiques, comme l’endroit de nidification, le niveau de spécialisation et la capacité d’être kleptoparasite, peuvent aider à comprendre comment se forment et s’assemblent les communautés (Forrest et al. 2015). De plus, ils permettent d’identifier les facteurs clés retrouvés dans l’environnement qui expliquent cet ensemble de traits et d’espèces. En effet, certaines espèces qui possèdent des traits mal adaptés à certains changements seront désavantagées comparativement aux autres espèces avec des traits plus avantageux (Rader et al. 2014; Persson et al. 2015; Arbetman et al. 2017). Parmi les espèces retrouvées au Québec, certaines se retrouvent en difficulté avec les conditions environnementales actuelles et parmi celles-ci, seulement trois ont été évaluées par le COSEPAC (Comité sur la situation des espèces en péril du Canada), soit Bombus affinis (en voie de disparition), Bombus bohemicus (en voie de disparition) et Bombus terricola (préoccupante) (Gouvernement du Canada 2016). Le déclin de ces espèces s’inscrit dans une décroissance populationnelle mondiale des pollinisateurs indigènes. Effectivement, plusieurs articles mettent en évidence la situation alarmante des bourdons en Amérique et en Europe (voir paragraphe suivant). Ce déclin ne semble pas toucher les espèces aléatoirement et un lien est à faire entre ce déclin et certains traits fonctionnels (Rader et al. 2014; Persson et al. 2015; Arbetman et al. 2017).
1.3. Déclin des pollinisateurs et principaux facteurs
Plusieurs espèces de pollinisateurs, dont les bourdons, ont montré d’importantes modifications dans leur distribution en Amérique du Nord, particulièrement B. occidentalis, B. pensylvanicus, B. affinis et B. terricola (Cameron et al. 2011; Bommarco et al. 2012; Kerr et al. 2015; Jacobson et al. 2018). En effet, leur abondance a chuté depuis les 50 dernières années et certaines espèces, comme B. affinis et B. bohemicus, ont même complètement disparu de certaines portions de leur aire de distribution (Kerr et al. 2015; Jacobson et al. 2018). Ces observations concordent avec la diminution d’abondance et de richesse documentée chez d’autres groupes de pollinisateurs (Potts et al. 2010; Durant and Otto 2019; Powney
et al. 2019). Le cheptel mondial de l’abeille à miel, par exemple, diminue d’année en année, et ce, malgré les multiples initiatives citoyennes, provinciales, nationales ou internationales qui tentent d’améliorer leurs conditions (Smith et al. 2013). D’autres groupes d’abeilles voient aussi leur abondance diminuer, notamment les espèces qui possèdent les plus petites niches écologiques (Rader et al. 2014; Powney et al. 2019). Ces pertes d’abondance ou d’espèces sont particulièrement inquiétantes et souvent causées indirectement ou directement par les humains. Un facteur important est sans aucun doute l’intensification agricole observée mondialement.
1.3.1. Intensification agricole
La perte, la fragmentation et la dégradation des habitats naturels, l’homogénéisation du paysage, entre autres par une moins grande diversité de cultures et une simplification des rotations de cultures, l’utilisation intensive de fertilisants et de pesticides (insecticides, fongicides et herbicides) par les régies de culture conventionnelle ou biologique, en plus du travail mécanique important du sol, sont tous des constituants ou des effets du concept général qu’est l’intensification agricole (Stoate et al. 2001; Benton et al. 2003; Tscharntke et al. 2005). Deux principales notions peuvent regrouper tous ces facteurs : la régie de culture et le paysage.
1.3.1.1 Régies de culture
Un nombre important d’études dans divers types de cultures a montré que la biodiversité des pollinisateurs indigènes, et particulièrement celle des bourdons, est influencée par la régie de culture qui est soit biologique ou conventionnelle. La différence majeure entre ces deux types de régies réside probablement dans leur gestion des pesticides, notamment celle des herbicides. De fait, les fermes biologiques sont soumises à un plus grand nombre de restrictions quant à leur usage des produits phytosanitaires. À ce titre, les produits phytosanitaires utilisés en régie conventionnelle, dont les pratiques agricoles sont souvent plus intensives, influenceraient négativement la présence des pollinisateurs retrouvée dans les cultures de canola (Morandin and Winston 2005), de blé (Clough et al. 2007; Gabriel
et al. 2013; Adhikari et al. 2019), des friches (Holzschuh et al. 2008) et des pâturages (Power et al. 2012; Power et al. 2016). Une étude qui recoupe cinq types de cultures dans quatre pays européens a montré que l’intensification de l’agriculture affecte négativement la diversité des pollinisateurs (Bartomeus et al. 2014). Cette différence en diversité ne se répercutait cependant pas sur le nombre de visites faites par les pollinisateurs sur les cultures; visites qui étaient probablement effectuées par les espèces dominantes du milieu (Bartomeus et al. 2014).
L’implication des pesticides dans le déclin des pollinisateurs est de plus en plus évidente (Blacquière et al. 2012; Rundlöf et al. 2015; Samson-Robert et al. 2015; Stanley et al. 2015a). De fait, les pollinisateurs sont exposés à un large cocktail de pesticides, notamment dans les zones cultivées selon une régie conventionnelle. En plus d’être présents dans l’habitat des pollinisateurs, insecticides, fongicides et herbicides sont retrouvés à des concentrations importantes (ex. thiaclopride : 2.9ng/g) dans l’alimentation des pollinisateurs via la consommation de pollen (David et al. 2016). Parmi les pesticides utilisés, on trouve une classe d’insecticides introduite au début des années 1990, les néonicotinoïdes, qui serait particulièrement nocive pour les insectes pollinisateurs (Feltham et al. 2014; Gill and Raine 2014; Stanley et al. 2015a). Cette classe de pesticides est largement utilisée en Amérique du Nord depuis la fin des années 2000. Leur stabilité et leur solubilité dans l’eau fait en sorte qu’ils sont très persistants dans l’environnement (DiBartolomeis et al. 2019; Giroux 2019; Montiel-León et al. 2019). En affectant le système nerveux des insectes, les néonicotinoïdes provoquent ultimement leur mort (Blacquière et al. 2012; Moffat et al. 2015; Stanley et al. 2015b; Siviter et al. 2018). Il s’agit de pesticides à large spectre qui affectent les insectes ciblés ou non par les traitements phytosanitaires. Les néonicotinoïdes les plus communs sont la clothianidine, le thiaméthoxame et l’imidaclopride. Leur impact sur les bourdons en laboratoire a montré des effets létaux et sous-létaux (Moffat et al. 2015), et leurs effets en milieu naturel semblent importants (Gill and Raine 2014; Kenna et al. 2019; mais voir Cutler and Scott-Dupree 2014).
Les pesticides ont des effets néfastes sur les ouvrières bourdons. Samson-Robert et al. (2015) ont utilisé l’accumulation de l’acétylcholinestérase comme proxy de la présence de pesticides néonicotinoïdes. L’acétylcholinestérase catabolise l’acétylcholine (neurotransmetteur) de son site actif. Cependant, le néonicotinoïde se lie au site actif à la place de l’acétylcholine. Cette réaction provoque un surplus de stimuli que le cerveau tente de contrôler en produisant un surplus d’acétylcholinestérases (Tomizawa and Casida 2001; Tomizawa 2013). Néanmoins, cette dernière ne peut pas déloger le pesticide, elle s’accumule donc au cerveau. Samson-Robert et al. (2015) a donc quantifié cette accumulation pour la mettre en relation avec la concentration de pesticides dans l’habitat des bourdons. Les colonies installées sur les parcelles d’agriculture intensive possédaient un niveau significativement plus important d’acétylcholinestérase que celles installées sur les parcelles en milieu extensif (production sans enrobage), montrant ainsi l’effet nocif des néonicotinoïdes au niveau du cerveau des bourdons. Les ouvrières intoxiquées aux néonicotinoïdes présentent également des signes de désorientation, de pertes de mémoire et de perte d’efficacité lors de leur quête alimentaire (Gill and Raine 2014; Moffat et al. 2015; Stanley et al. 2015b; Siviter et al. 2018; Kenna et al. 2019).
L’effet des pesticides sur les ouvrières se reflète également sur la qualité de la pollinisation effectuée par les bourdons. Une étude effectuée dans la culture de la pomme a montré que les néonicotinoïdes affectent négativement la qualité de la pollinisation fournie par les bourdons (Stanley et al. 2015a). En effet, les colonies en contact avec le pesticide fréquentaient moins souvent et moins efficacement les fleurs de pommiers, avec pour conséquence une production de pommes de moins bonne qualité (Stanley et al. 2015a). La diminution de la qualité de pollinisation causée par les pesticides a en plus été constatée pour d’autres cultures comme le canola, le tournesol et le melon (Challa et al. 2019; Tschoeke et al. 2019; de Oliveira et al. 2019). Les pesticides affectent également le développement et la reproduction des colonies. En effet, des tests faits en milieu naturel et en laboratoire arrivent presque tous aux mêmes conclusions : l’initiation des colonies est plus difficile, elles prennent moins de poids et produisent généralement moins d’ouvrières, de mâles
et de reines en présence de pesticides (Gill et al. 2012; Whitehorn et al. 2012; Goulson 2015; Stanley et al. 2016; Baron et al. 2017; Ellis et al. 2017; Stanley and Raine 2017; Wu-Smart and Spivak 2018). Ces études n’ont cependant pas mesuré la longévité ainsi que les stratégies de butinage des colonies. En effet, elles ont plutôt évalué le nombre d’ouvrières produites en sacrifiant les colonies à un moment déterminé de leur développement.
Les fongicides ne visent pas directement les pollinisateurs indigènes, mais leurs effets néfastes, lorsque combinés aux insecticides, sont de plus en plus mis en évidence. La présence des fongicides dans le cocktail de pesticides augmente la toxicité des insecticides et agit ainsi en synergie contre les pollinisateurs (David et al. 2016). En effet, le taux de mortalité des ouvrières de Bombus terrestris était significativement plus important dans les traitements combinant fongicides et insecticides (à l’exception de l’imidaclopride) contrairement aux traitements comprenant le fongicide ou l’insecticide seul (Raimets et al. 2018). Même l’exposition seule à des fongicides chez Bombus impatiens a influencé négativement la production d’ouvrières, la masse totale prise par la colonie ainsi que la masse des nouvelles reines produites (Bernauer et al. 2015).
Bien que les herbicides utilisés par les agriculteurs ne semblent pas affecter directement la santé des bourdons, ils ont néanmoins un impact sur leur survie (Potts et al. 2010). De fait, les bourdons sont habituellement généralistes, c’est-à-dire qu’ils vont habituellement se nourrir de plusieurs espèces de fleurs. Pour se développer de manière efficace et combler tous leurs besoins en acides aminés essentiels, ils vont utiliser différentes sources de pollen. La diminution de la diversité florale sur les fermes, soit par l’utilisation d’herbicides ou tout simplement par la tonte des bordures de champs, affecte négativement la survie des pollinisateurs indigènes (Bretagnolle and Gaba 2015; Vaudo et al. 2016; Vaudo et al. 2018). Des études ont montré qu’en plantant différentes espèces de fleurs en bordure de champs agricoles, la diversité des pollinisateurs indigènes et la production du champ en question
étaient significativement augmentées (Blaauw and Isaacs 2014a; Blaauw and Isaacs 2014b).
1.3.1.2. Paysage
Plusieurs études ont tenté de montrer l’effet de la composition, de la configuration et de l’hétérogénéité du paysage sur les pollinisateurs indigènes, particulièrement les bourdons. La composition du paysage fait référence aux types de parcelles d’habitat présentes (culture intensive, forêt, pâturage, etc.) dans un rayon déterminé ainsi qu’à leurs couverts relatifs. La configuration du paysage, quant à elle, concerne aussi bien les caractéristiques topographiques d’un ensemble de parcelles (exemples : forme, taille et localisation) que les relations spatiales avec d’autres éléments du paysage (exemples : haies brise-vent, bandes riveraines et bordures des champs) (Tableau 1 in Xiao et al. 2016).
La composition du paysage a un impact considérable sur les bourdons, autant pour les ouvrières que pour les colonies. En effet, un paysage qui contient une importante proportion de parcelles fleuries aura nécessairement un impact sur la quête alimentaire des pollinisateurs. À ce titre, une étude a quantifié la distance de vol (du nid à la source florale) de bourdons selon la composition et la configuration du paysage (Redhead et al. 2016). La distance de vol du nid vers les parcelles de nourriture était nettement plus faible pour les ouvrières qui habitaient des paysages composés de parcelles plus diversifiés ayant un couvert floral important (Redhead et al. 2016). Bien que les auteurs n’aient pu comparer le développement des différentes colonies, les effets associés à un vol court des ouvrières ont certainement des répercussions bénéfiques sur les colonies. De manière similaire, les colonies placées dans des paysages contenant plus de ressources florales ont pris plus de poids et produit plus d’ouvrières et de mâles que les colonies placées dans les paysages avec une moins grande proportion d’habitats fleuris (Williams et al. 2012; Spiesman et al. 2017). Le type d’habitats présents dans le paysage affecte également le développement des colonies. Les colonies présentes dans les milieux semi-naturels ou naturels avaient, par exemple, produit plus de reines que les
colonies dans les milieux agricoles conventionnels ou en banlieues (Crone and Williams 2016; Milano et al. 2019).
La composition du paysage affecte également les communautés d’abeilles indigènes. Plus les proportions d’habitats naturels ou semi-naturels sont importantes, plus l’abondance et la richesse des pollinisateurs, ainsi que l’efficacité de leur pollinisation, le sont également (Kremen et al. 2002; Kremen et al. 2004; Greenleaf and Kremen 2006; Le Féon et al. 2010). En effet, lorsque plus de 30% d’habitats naturels ou semi-naturels étaient conservés dans un rayon de 2 km autour des champs, l’abondance et la richesse spécifiques des abeilles augmentaient significativement (Kremen et al. 2002). La présence d’habitats naturels à proximité des cultures permet également de diminuer l’impact négatif des pesticides sur les communautés de pollinisateurs (Park et al. 2015). En effet, la sensibilité aux pesticides des pollinisateurs indigènes (mesurée en termes d’abondance et de richesse spécifique) était dépendante de la quantité d’habitats naturels (%) dans le paysage environnant (Park et al. 2015). Ceci dit, certains habitats non naturels, comme les prairies, sont favorables à la richesse spécifique et à l’abondance des pollinisateurs puisqu’ils leur offrent des ressources nidicoles et alimentaires de meilleure qualité (Sjödin et al. 2008; Kennedy et al. 2013; Bennett and Isaacs 2014; Martins et al. 2018).
L’effet de la configuration du paysage sur les bourdons semble de dépendre du proxy utilisé pour le quantifier. Au niveau de l’impact sur les ouvrières, une étude suédoise a montré que les ouvrières qui évoluent dans des paysages plus complexes, et ce, tant au niveau de la composition que de la configuration, possédaient une taille significativement plus grande (Persson and Smith 2011). Selon les auteurs, la disponibilité des ressources serait davantage limitée dans les paysages moins complexes, ce qui pourrait entrainer de la compétition et diminuer la capacité de la colonie à produire des ouvrières de plus grande taille. Ces ouvrières, moins efficaces à amasser les ressources, affecteraient également le développement même de la colonie. La complexité du paysage affecte aussi la
distance qu’une ouvrière doit parcourir pour trouver de la nourriture. Plus un paysage est diversifié et possède une grande quantité de ressources florales, plus la distance de vol requise pour y accéder à partir du nid risque d’être petite (Redhead et al. 2016). Ceci implique que dans les paysages moins complexes et moins riches en fleurs, les bourdons devraient voyager plus loin en moyenne, ce qui affecterait négativement la dynamique de la colonie. À l’opposé, lorsque le proxy de la configuration du paysage mesuré était la connectivité, les colonies de bourdons ne semblaient pas influencées, puisque les colonies en milieu isolé avaient pris la même quantité de poids que les colonies en milieu « connecté » (Herrmann et al. 2017). Au niveau des communautés, la quantité de routes ou de bordures (incluant bandes riveraines et haies brise-vent) semble influencer positivement l’abondance et la richesse des pollinisateurs (Sjödin et al. 2008; Martins et al. 2015). À l’inverse, la configuration du paysage en utilisant trois types d’index (ratio aire/périmètre, plus près voisin ainsi qu’interjection et juxtaposition) n’a pas influencé la richesse et l’abondance des pollinisateurs indigènes étudiés (Kennedy et al. 2013). L’impact des différents proxys de la configuration du paysage sur les bourdons gagnerait à être testé, puisqu’aucun consensus n’existe actuellement.
1.3.2. Autres facteurs dans le déclin des bourdons 1.3.2.1. Changements climatiques
Les changements climatiques impliquent une modification durable des paramètres statistiques comme les températures moyennes, les précipitations moyennes et les extrêmes de température dans le climat mondial ou régional (Bush and Lemmen 2019). Phénomène habituellement naturel, il est dangereusement amplifié par les humains depuis la révolution industrielle (Bush and Lemmen 2019). Dans le Sud du Québec, les températures observées, depuis les cinq dernières décennies, ont augmenté de 1 à 3°C. Or, les températures annuelles devraient encore augmenter d’environ 2 à 4 °C à l’horizon 2070, et de 4 à 7 °C pour la période 2070-2100 (Bush and Lemmen 2019). Ces augmentations, même de quelques degrés, ont des conséquences considérables sur les écosystèmes.
Les principales réponses écologiques connues des organismes envers les changements climatiques sont le décalage de l’aire de distribution des espèces vers les pôles(Lancaster et al. 2015), le changement de la phénologie du cycle vital des espèces (Durant et al. 2007) et la diminution de la taille des espèces et des individus, principalement chez les ectothermes (Gardner et al. 2011; Sheridan and Bickford 2011). Contrairement à la majorité des organismes étudiés, l’aire de distribution des bourdons de l’hémisphère nord ne semble pas se déplacer vers le nord. En effet, une étude a utilisé des données d’occurrence de bourdons des 110 dernières années pour l’Amérique du Nord et l’Europe (Kerr et al. 2015). Leurs conclusions : les 67 espèces de bourdons étudiés d’un continent à l’autre partagent toutes la même réponse aux changements climatiques, soit une diminution de leur présence dans le Sud et un statu quo de leur présence dans le Nord. Leur aire de distribution se voit donc de plus en plus réduite au fil des années (Kerr et al. 2015). De plus, l’abondance de plusieurs bourdons, dont Bombus affinis (passé de 1859 spécimens capturés en 1974 à 74 en 2010), B. bohemicus (passé de 530 spécimens capturés en 1974 à 80 en 2010) et B. borealis (passé de 708 spécimens capturés en 1974 à 153 en 2010) a drastiquement diminué en Amérique du Nord.
Les bourdons, comme tout organisme, sont dépendants de leur source de nourriture et de leur environnement. Les changements de température enregistrés dans les dernières années ont eu pour effet de modifier à la fois la phénologie de leurs sources de nourriture en plus de leur propre phénologie (Forrest 2015; Miller-Struttmann et al. 2015). En conséquence, des décalages phénologiques entre les plantes et leurs pollinisateurs spécialistes sont de plus en plus observés (Forrest 2015). En Pennsylvanie, des données historiques sur la présence de deux bourdons spécialistes (B. balteatus et B. sylvicola) en région alpine ont permis de montrer un changement dans leur régime alimentaire (Miller-Struttmann et al. 2015). Au départ, ces espèces à glosse longues étaient principalement adaptées à la présence de fleurs à longue corolle. Au fil du temps, et dû aux changements climatiques, d’autres espèces de bourdons ont rejoint ces régions alpines et ont fait concurrence aux deux
bourdons natifs. Ces derniers, pour pouvoir survivre à cette compétition interspécifique, ont dû élargir leur niche alimentaire afin d’accéder à plusieurs types de fleurs. Ceci a été permis par une réduction graduelle de la longueur de leur glosse (Miller-Struttmann et al. 2015). Une autre étude a également montré que l’augmentation prévue des températures pouvait affecter la phénologie des abeilles. Suite à des tests effectués en laboratoire, ils ont observé que l’abeille Plebeia droryana (Friese) (Hymenoptera: Apidae : Meliponini), une espèce qui vit au Brésil et dont la diapause reproductive est connue, perd sa diapause avec l’augmentation des températures (Dos Santos et al. 2015). Ce changement dans le cycle de vie de cette espèce pourrait grandement affecter sa niche écologique et les espèces de plantes qu’elle pollinise. Les changements climatiques affectent donc la phénologie des insectes en plus des relations trophiques interspécifiques. Ces conséquences pourraient donc grandement affecter les espèces spécialistes menacées ou en péril.
1.3.2.2. Maladies, parasites et prédateurs
Plusieurs organismes vivants vont nuire à la santé et à la survie des colonies de bourdons, autant chez les colonies commerciales que les colonies sauvages. Ces organismes peuvent se retrouver naturellement dans l’environnement ou provenir d’hôtes différents, comme l’abeille à miel. Certains parasites vont pouvoir être transmis des abeilles à miel aux bourdons sauvages, et similairement, les bourdons commerciaux vont pouvoir transmettre leurs parasites aux pollinisateurs indigènes. En effet, des bourdons sauvages retrouvés à proximité (< 500 m) de ruches d’abeilles à miel ou de colonies de bourdons commerciales présentaient des prévalences en parasites (Apicytis bombi et Crithidia bombi) beaucoup plus élevés que les colonies placées à 5 km (~18%; Graystock, Goulson, & Hughes, 2014). La transmission de maladies et parasites aux populations de bourdons indigènes par le biais de pollinisateurs commerciaux inquiète de plus en plus et compte parmi les causes les plus citées du déclin des pollinisateurs. Les infections fongiques Nosema bombi et Crithidia bombi, certains virus transmis par les acariens, ainsi que les papillons qui se spécialisent dans la destruction des nids, comme la pyrale du bourdon, Aphomia sociella (Lepidoptera : Pyralidae), sont tous des parasites ou
prédateurs qui peuvent grandement nuire au développement des colonies de bourdons.
Nosema bombi et C. bombi sont des parasites obligatoires intracellulaires faisant partie du règne des champignons. Ils se retrouvent dans le système digestif des bourdons et sont généralement inoffensifs (Imhoof and Schmid-Hempel 1999), mais peuvent causer d’importants dommages lorsque la reine et les colonies sont en période de stress. Lorsque testées en milieu contrôlé, les colonies infectées par N. bombi ont produit moins de mâles et pratiquement aucune reine (Otti and Schmid-Hempel 2007). Similairement, lorsque les colonies infectées à C. bombi étaient affamées, leur succès reproducteur diminuait de près de 40% et la durée de vie de leurs ouvrières était réduite en moyenne de 50% (Brown et al. 2000; Brown et al. 2003).
Certains virus associés à l’abeille à miel, comme le virus de l’aile déformée (DWV), peuvent également toucher les populations de bourdons sauvages. Bien qu’ils soient transmis à l’abeille à miel par le biais de la mite Varroa destructor, la voie orale (via le pollen et le nectar des fleurs) semble être celle privilégiée pour contaminer les populations de bourdons indigènes. En effet, même en l’absence du varroa, les bourdons indigènes sont tout de même infectés (Genersch et al. 2006). Ces virus sont parfois même plus virulents chez le bourdon, puisqu’un changement d’hôtes entraine souvent un changement dans la virulence du virus (Fürst et al. 2014).
Un autre ennemi des bourdons est la pyrale des bourdons, A. sociella. Ce papillon se nourrit, lors de ses stades larvaires, de toutes les composantes du nid des bourdons, soit la cire, le pollen, le nectar et le couvain. Les larves se développent et peuvent complètement saccager un nid de bourdons mature sur une période d’environ quarante jours (Pouvreau 1988). La valeur adaptative des colonies n’est pas trop réduite, puisque les chenilles attaquent le nid souvent après le début de la production des gynes (futures reines) (Goulson et al. 2018). Les infestations peuvent