Nétrine et cellules souches, attirance ou répulsion ?
Texte intégral
Figure
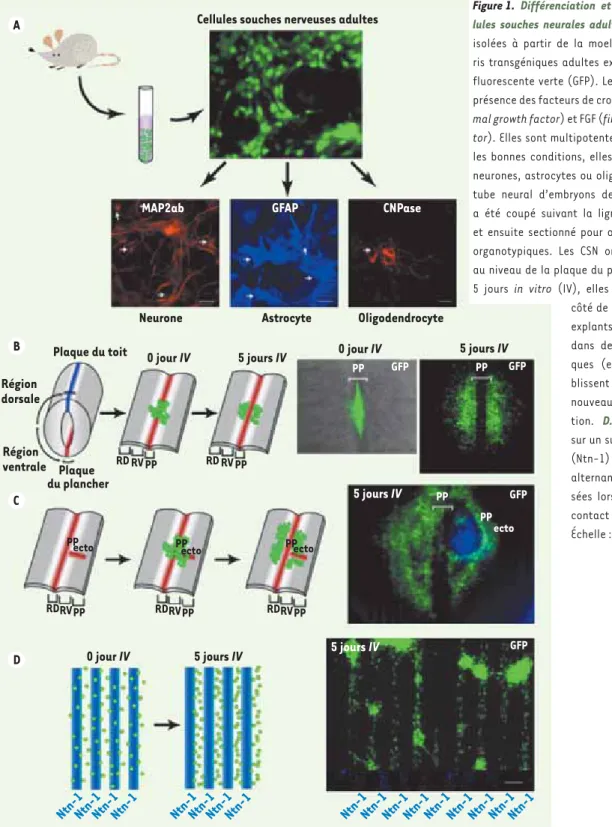
Documents relatifs
Il est exprimé dans les cellules progénitrices en division et les neurones granulaires im- matures dans le gyrus denté adulte mais pas dans les cellules progénitrices de
Pour répondre à ces objectifs, notre projet se déployait en quatre grands volets complémentaires, les deux premiers (documentation et
Près de 44 000 cas humains ont été déclarés à l’OMS par 26 pays au cours des vingt derniè- res années (Tableau I) [1, 2] , et le nombre réel de cas est probablement
Under clear-sky conditions, all 19 models that provide clear-sky fluxes show an overall decrease (statistically sig- nificant at the 95% level), but again of largely varying
147 In the zebrafish, I demonstrated that the adult pallial germinal zone derives from the two different progenitor populations: the embryonic “proneural
Pour cela, les cellules de faible densité de la moelle de souris ont été d'abord recueillies sur gradient de densité, puis les cellules différenciées ont été
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
As expected, the profound changes of the HIV-1 splicing pattern caused by the overexpression of one of the SR pro- teins ASF/SF2, SC35 or 9G8 inhibited viral protein synthe-
