Mechanisms and PK/PD modelling of MCR-1-induced adaptive resistance in Enterobacteriaceae
Texte intégral
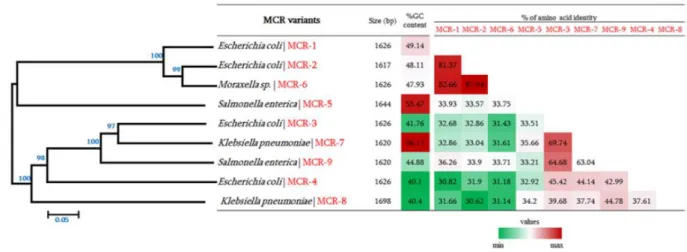
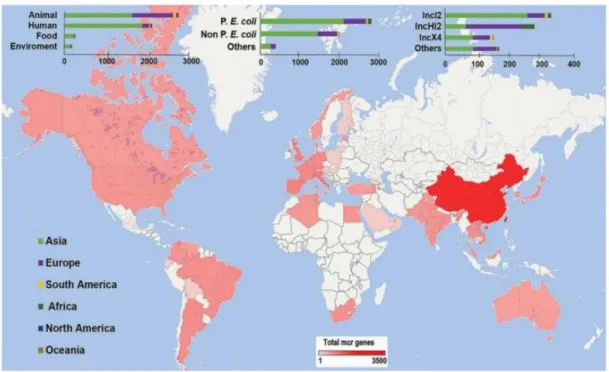
Figure
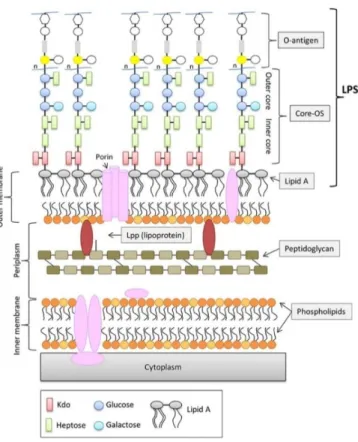
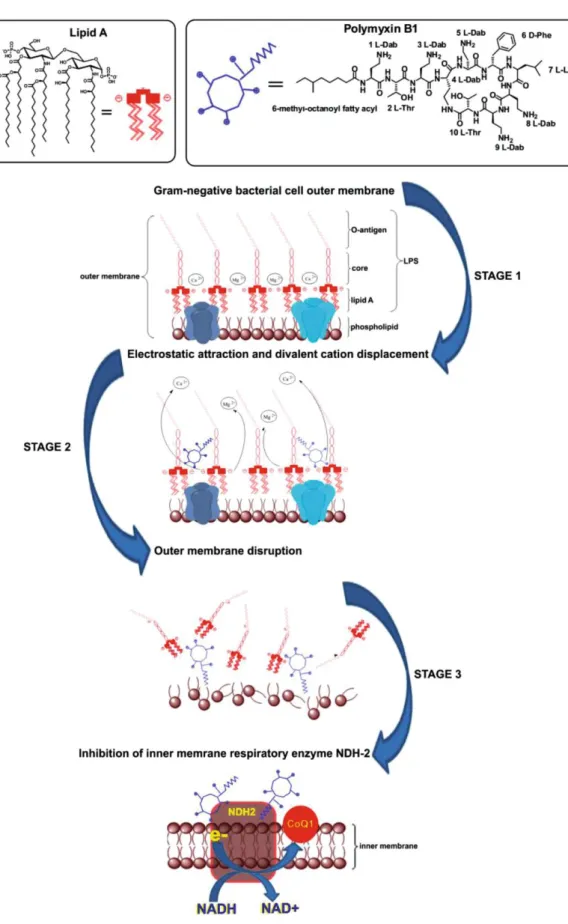
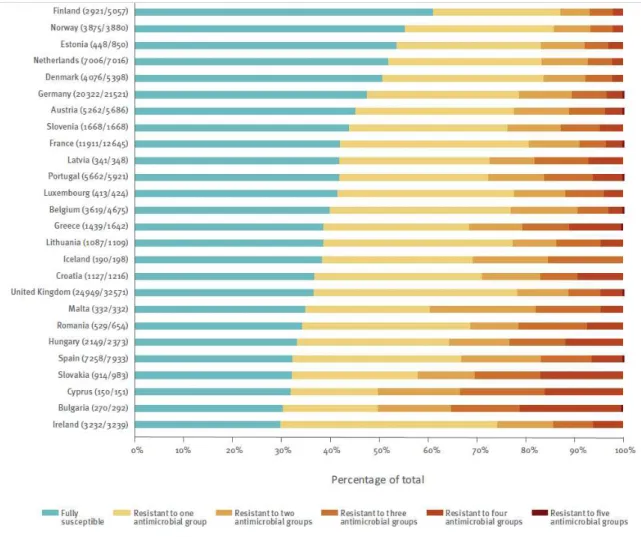
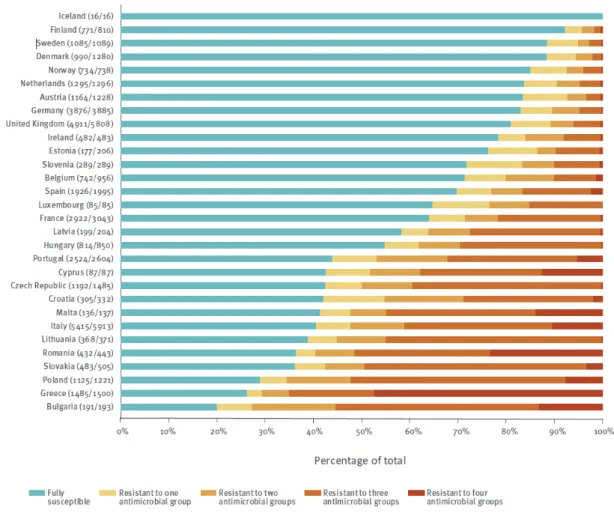
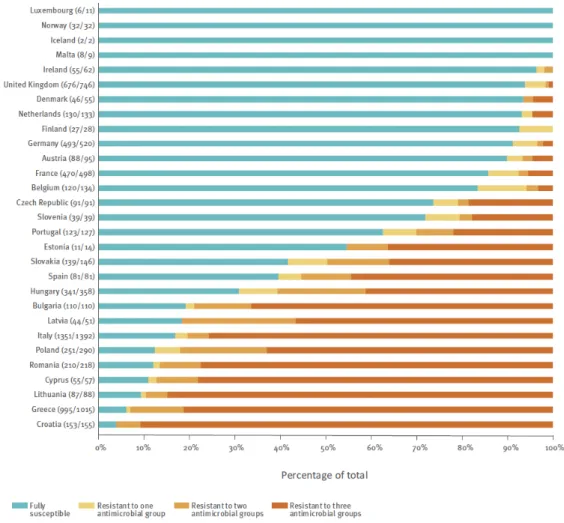
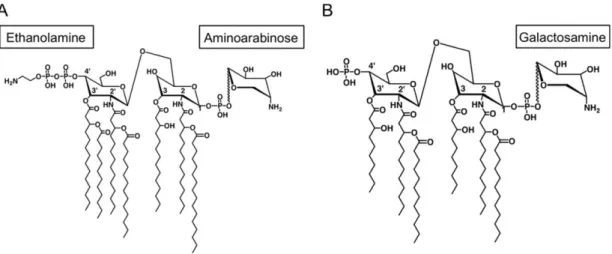
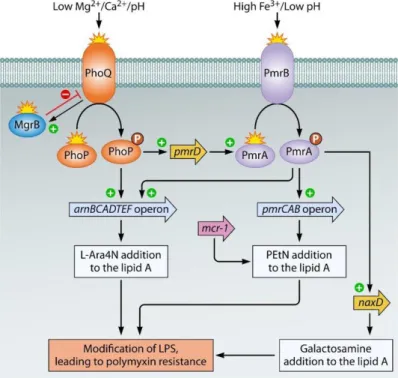
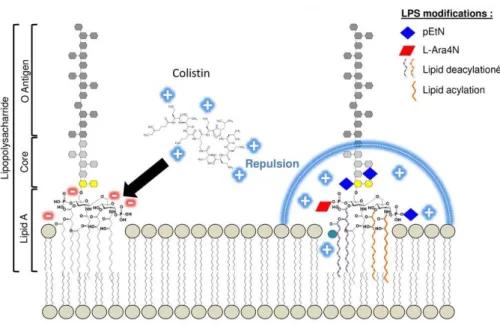
Documents relatifs
Bacterial genes also allow the synthesis of vitamins ( Yoshii et al., 2019 ), neurotransmitters ( Onaolapo et al., 2020 ), and metabolites derived from tryptophan ( Agus et al., 2018
Conversely, a diet en- riched with 20% rapeseed oil results in a lower inflammation both in the circulation and in the adipose tissue, notably via an increase of plasma
[r]
[r]
Ab dem Durchbruch der Zähne vollzieht sich eine langsame aber stetige Einengung der Pulpa, was letztendlich zu einer voll- ständigen Obliteration der Pulpenkam- mer führt..
In a [ 13 C, 1 H]- HSQC spectrum recorded at a proton resonance fre- quency of 900 MHz (Figure 4), most of the peaks in the crowded spectral regions could readily be identi- fied,
Additionally, the Monte Carlo simulations involving the angular distributions of x-rays demonstrate that the proposed approach describes well the RR enhancement measured in
